
산화적 인산화(酸化的 燐酸化, 영어: oxidative phosphorylation)는 전자전달과 화학삼투를 통한 ATP의 합성과정이다. 대부분의 진핵생물에서 산화적 인산화는 미토콘드리아에서 일어난다. 거의 모든 산소 호흡을 하는 생물들은 산화적 인산화를 수행한다. 산화적 인산화는 혐기적인 발효 과정과 비교했을 때 호흡 기질이 분해될 때 방출되는 에너지로 ATP를 합성하는 매우 효율적인 방법이기 때문에 자연계에 널리 펴져있다.
산화적 인산화가 일어나는 동안 전자는 산화환원반응을 통해 전자 공여체로부터 산소(O2)와 같은 전자 수용체로 전달된다. 이러한 산화환원반응으로 방출된 에너지는 ATP를 합성하는데 사용된다. 진핵생물에서 이러한 산화환원반응은 미토콘드리아 내막에 있는 일련의 단백질 복합체들에 의해 수행된다. 원핵생물에서는 이러한 단백질들이 세포막에 존재한다. 이렇게 연결된 일련의 단백질들의 세트를 전자전달계라고 부른다. 진핵생물에서는 5가지 주요 단백질 복합체가 관여하는 반면에 원핵생물에서는 여러 가지 전자 공여체와 전자 수용체를 사용하는 다양한 효소들이 존재한다.
이러한 전자전달계를 통해 전달되는 전자에 의해 방출된 에너지는 미토콘드리아 기질에서 막 사이 공간으로 H+(양성자)를 능동수송하는데 사용된다. 이것은 미토콘드리아 내막을 경계로 H+의 농도 기울기와 전위 차이의 형태로 위치 에너지를 발생시킨다. 미토콘드리아 내막을 경계로 생성된 H+의 전기화학적 기울기에 의해 H+가 ATP 생성효소를 통해 막 사이 공간에서 미토콘드리아 기질로 확산될 때 ATP가 합성되고, 이 과정을 화학삼투라고 한다. ATP 생성효소는 ADP를 ATP로 전환시키는 인산화 반응에 에너지를 사용한다. 이 과정에서 ATP 생성효소의 일부가 회전하는데 이 힘은 H+의 확산에 의해 제공된다.
산화적 인산화는 물질대사의 중요한 부분이지만, 초과산화물 및 과산화 수소와 같은 활성산소를 생성하여 자유 라디칼의 증가, 세포 손상을 유도하고, 질병 및 노화에도 영향을 미친다. 또한 산화적 인산화를 수행하는 효소들은 그 활동을 저해하는 많은 약물과 독소들의 표적이다.
산화적 인산화는 진핵생물에서 세포 호흡의 마지막 과정이며 높은 ATP 수율을 가지고 있다.
화학삼투
산화적 인산화는 에너지 방출 반응을 사용하여 에너지 흡수 반응을 진행시킨다. 두 세트의 반응이 짝지어져 있다고 하는데, 이는 두 반응 중 하나가 작동하지 않으면 다른 하나도 작동할 수 없다는 것을 의미한다. NADH와 같은 전자 공여체에서부터 산소(O2)와 같은 전자 수용체에 이르기까지 전자전달계를 통한 전자의 흐름은 에너지를 방출하는 과정이다. 반면에 ATP 합성은 에너지를 흡수하는 과정이다. 전자전달계와 ATP 생성효소는 모두 미토콘드리아 내막에 존재하고, 미토콘드리아 내막을 통과하는 H+의 움직임에 의해 전자전달계에서 ATP 생성효소로 에너지가 전달되는데, 이러한 과정을 화학삼투라고 한다. 실제로 이것은 단순한 전기 회로 같으며, H+는 전자전달계의 H+ 펌핑 효소들에 의해 미토콘드리아 기질(N쪽, 상대적으로 H+ 농도가 낮은 쪽)에서 막 사이 공간(P쪽, 상대적으로 H+ 농도가 높은 쪽)으로 능동수송된다. 이러한 효소들은 배터리와 같으며, 회로를 통해 전류를 흐르게 하는 일을 수행한다. H+(양성자)의 움직임은 막을 가로질러 전기화학적 기울기를 만들어 내는데, 이것은 종종 양성자 구동력이라고 불린다. 양성자 구동력은 H+의 농도 차이(양성자 기울기, ΔpH)로 인한 화학적인 위치 에너지와 H+가 상대 이온 없이 막을 지나 이동할 때, 전하의 분리 차이로 인한 전기적인 위치 에너지의 2가지 구성 요소를 갖는다.
H+가 전기화학적 기울기에 따라 막 사이 공간(P쪽)에서 미토콘드리아 기질(N쪽)로 ATP 생성효소를 통해 확산될 때 방출되는 에너지로 ATP를 합성한다. 이러한 전기화학적 기울기는 ATP 생성효소의 일부를 회전시키는 구동력으로 작용하고, 이러한 움직임은 ATP 합성과 짝지워진다.
양성자 구동력의 2가지 구성 요소는 열역학적으로 동등하다. 미토콘드리아에서 에너지의 가장 큰 부분은 전위차에 의해 주어진다. 반면에 엽록체는 주로 ΔpH 차이에 의해 작동한다.
산화적 인산화에 의해 방출되는 에너지의 양은 혐기성 발효에 의해 생성되는 에너지의 양보다 크다. 해당과정은 포도당 1분자당 2ATP를 생산하지만, 산화적 인산화는 포도당 1분자당 28~34ATP를 생산한다.지방산의 β 산화는 각 사이클당 약 14ATP를 생산한다. 이러한 ATP의 수율은 이론적인 최대값이다. 실제로 일부 H+(양성자)는 막을 통해 누출되기 때문에, ATP의 수율을 떨어뜨린다.
전자 및 양성자 전달 분자
전자전달계는 양성자와 전자를 모두 운반하고, 전자를 전자 공여체에서 전자 수용체로 전달하고, H+(양성자)를 막을 가로질러 능동수송한다. 이 과정에서 가용성 전달 분자 및 단백질에 결합된 전달 분자를 사용한다. 미토콘드리아에서 전자는 수용성 전자전달 단백질인 사이토크롬 c에 의해 막 사이 공간 내에서 전달된다. 전자전달 단백질 내의 헴기가 가지고 있는 철 원자의 산화와 환원에 의해 전자가 전달된다. 사이토크롬 c는 몇몇 세균의 주변세포질 공간에서도 발견된다.
미토콘드리아 내막에 존재하는 지용성 전자운반체인 유비퀴논(조효소 Q10 또는 Q)은 산화환원 회로에 의해 전자와 H+(양성자)를 둘 다 운반한다. 유비퀴논은 작은 벤조퀴논 분자로 매우 소수성이어서 막 내부에서 자유롭게 확산된다. 유비퀴논(Q)이 2개의 전자와 2개의 H+를 받아들이면, 유비퀴놀(QH2)의 형태로 환원된다. 유비퀴놀(QH2)이 2개의 전자와 2개의 H+를 방출하면, 산화되어 다시 유비퀴논(Q)의 형태로 되돌아간다. 유비퀴논(Q)이 막의 한 쪽에서 환원되고, 다른 한 쪽에서 유비퀴놀(QH2)이 산화되면, 이러한 반응들을 결합시켜 막을 통해 H+를 능동수송한다. 일부 세균의 전자전달계는 유비퀴논 외에 메나퀴논과 같은 다른 퀴논을 사용한다.
단백질 내에서 전자는 플라빈 보조 인자, 철-황 클러스터 및 사이토크롬 사이에서 전달된다. 철-황 클러스터에는 몇 가지 유형이 있다. 전자전달계에서 발견되는 가장 단순한 종류는 2개의 황 원자와 연결되어 있는 [2Fe-2S] 클러스터이다. 두 번째 종류인 [4Fe-4S]는 4개의 철 원자와 4개의 황 원자를 가진 정육면체 구조를 포함한다. 이들 클러스터의 철 원자는 일반적으로 시스테인의 황 원자와 배위 결합을 한다. 보조 인자인 금속 이온은 H+(양성자)와 결합하거나 또는 양성자를 방출하지 않고 산화환원반응을 거치기 때문에 전자전달계에서 단백질을 통해 전자를 전달하는 역할을 수행한다. 전자는 이러한 보조 인자의 사슬을 따라 단백질을 통해 도약함으로써 먼 거리를 이동한다. 이것은 1.4×10−9 m 미만의 거리에서 급격한 양자 터널링에 의해 발생한다.
진핵생물의 전자전달계
해당과정, 시트르산 회로, 지방산의 β 산화와 같은 많은 생화학적 이화작용들은 환원된 조효소인 NADH를 생성한다. NADH는 고에너지 전자를 가지고 있어서 산화될 때 많은 양의 에너지를 방출한다. 그러나 세포는 이 에너지를 한꺼번에 방출하지 않는다. 대신 NADH에서 전자를 떼어내어 일련의 전자전달효소들을 통해 전자를 산소로 전달하는 각 과정에서 소량의 에너지가 방출된다. 복합체 I~IV로 구성된 효소들의 세트는 전자전달계라고 불리며, 미토콘드리아 내막에 존재한다. 석신산도 전자전달계에서 산화되지만, 복합체 II를 통해 유입된다.
진핵생물에서 전자전달계의 효소는 NADH가 산화될 때 방출되는 에너지를 이용하여 미토콘드리아 기질에서 막 사이 공간으로 H+(양성자)를 능동수송한다. 이것은 막 사이 공간의 H+(양성자)의 농도를 증가시키고, 미토콘드리아 내막을 가로질러 전기화학적 기울기를 생성시킨다. 이러한 위치 에너지는 ATP 생성효소에 의해 ATP를 생성하는데 사용된다. 진핵생물의 미토콘드리아에서 일어나는 산화적 인산화는 이 과정에 대한 가장 잘 알려져 있는 예이다. 미토콘드리아는 Trichomonas vaginalis 와 같은 혐기성 원생동물을 제외하고 거의 모든 진핵생물에 존재한다. 대신에 Trichomonas vaginalis는 하이드로게노솜(hydrogenosome, 수소발생소포)라 불리는 미토콘드리아 잔존 세포소기관에서 H+(양성자)를 수소로 환원시킨다.
| 호흡 효소 | 산화환원 쌍 | 표준 환원 전위
(Volts) |
|---|---|---|
| NADH 탈수소효소 | NAD+ / NADH | −0.32 |
| 석신산 탈수소효소 | FMN or FAD / FMNH2 or FADH2 | −0.20 |
| 사이토크롬 bc1 복합체 | 유비퀴논산화형 / 유비퀴논환원형 | +0.06 |
| 사이토크롬 bc1 복합체 | 사이토크롬 b산화형 / 사이토크롬 b환원형 | +0.12 |
| 복합체 IV | 사이토크롬 c산화형 / 사이토크롬 c환원형 | +0.22 |
| 복합체 IV | 사이토크롬 a산화형 / 사이토크롬 a환원형 | +0.29 |
| 복합체 IV | O2 / HO− | +0.82 |
| 조건: pH = 7 | ||
NADH:유비퀴논 산화환원효소 (복합체 Ⅰ)
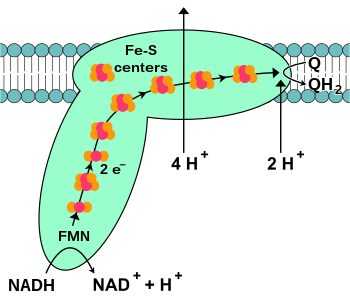
NADH:유비퀴논 산화환원효소 또는 NADH 탈수소효소 또는 복합체 I은 전자전달계의 첫 번째 단백질이다. 복합체 I은 46개의 소단위체로 구성된 분자량이 약 1,000 kDa 인 거대 효소이다. 세균에서 복합체 I의 구조는 자세하게 알려져 있다. 대부분의 생물에서 복합체 I은 큰 볼이 있는 부츠 모양을 닮았다. 미토콘드리아에 존재하는 많은 효소들의 경우와 마찬가지로 복합체 I을 구성하는 개별 단백질들을 암호화하는 유전자들은 세포핵과 미토콘드리아 게놈에 존재한다.
복합체 I에 의해 촉매되는 반응은 NADH의 2개의 전자를 유비퀴논(또는 조효소 Q10, 아래식에서 Q로 표시)으로 전달하는 산화반응이다. 유비퀴논은 미토콘드리아 내막에서 발견되는 지용성 퀴논이다.
-
()

전자전달계의 시작은 NADH가 복합체 I에 결합하여 2개의 전자를 제공하는 것이다. 전자는 복합체 I에 결합된 보결분자단인 플라빈 모노뉴클레오타이드(FMN)를 통해 복합체 I으로 들어간다. FMN이 전자를 얻으면 환원형인 FMNH2로 변환된다. 그런 다음 전자는 복합체 I의 두 번째 보결분자단인 일련의 철-황 클러스터를 통해 전달된다. 복합체 I에는 [2Fe–2S] 와 [4Fe–4S] 철-황 클러스터가 둘 다 존재한다.
전자가 복합체 I을 통과함에 따라 4개의 H+(양성자)가 미토콘드리아 기질에서 막 사이 공간으로 능동수송된다. 정확하게 이 과정에 어떻게 일어나는지는 분명하지 않지만, 복합체 I에서 구조적인 변화가 일어나서 단백질이 미토콘드리아 기질(N쪽)에서 H+(양성자)를 결합시키고, 막 사이 공간(P쪽)에서 H+(양성자)를 방출시키는 것으로 보인다. 마지막으로 전자는 철-황 클러스터 사슬에서 미토콘드리아 내막의 유비퀴논으로 전달된다. 유비퀴논의 환원은 H+(양성자)의 농도 기울기 생성에도 기여하는데, 유비퀴논(Q)이 유비퀴놀(QH2)로 환원됨에 따라 미토콘드리아 기질에서 2개의 H+(양성자)가 감소한다.
석신산 탈수소효소 (복합체 Ⅱ)

석신산 탈수소효소 또는 복합체 II 또는 석신산:유비퀴논 산화환원효소는 전자전달계의 두 번째 진입 지점이다. 석신산 탈수소효소는 시트르산 회로와 전자전달계에 둘 다 관여하는 유일한 효소이기 때문에 특이하다. 복합체 II는 보조 인자인 플라빈 아데닌 다이뉴클레오타이드(FAD), 철-황 클러스터, 헴기를 가지고 있는 4개의 단백질 소단위체로 구성된다. 복합체 II의 헴기는 유비퀴논으로 전자를 전달하진 않지만, 활성산소의 생성을 감소시키는데에 중요한 역할을 하는 것으로 여겨지고 있다. 복합체 II는 석신산을 푸마르산으로 산화시키고, 유비퀴논을 환원시킨다. 이 반응은 NADH의 산화보다 더 적은 에너지를 방출하기 때문에, 복합체 II는 막을 가로질러 H+(양성자)를 능동수송하지 않고, H+(양성자)의 농도 기울기 형성에 기여하지 않는다.
-
()

기생충인 돼지회충(Ascaris suum)과 같은 일부 진핵생물에서 복합체 II와 유사한 효소인 푸마르산 환원효소(메나퀴놀:푸마르산 산화환원효소)는 유비퀴놀을 산화시키고 푸마르산을 환원시키는 역반응을 촉매한다. 이것은 전자 수용체로 푸마르산을 사용하는 혐기성 산화적 인산화를 수행하여 기생충이 대장 내의 혐기성 환경에서 생존할 수 있도록 한다. 복합체 II의 또 다른 특이한 기능은 말라리아 기생충인 Plasmodium falciparum에서 찾아볼 수 있다. 여기서는 복합체 II가 산화효소로써 역반응에 관여하는데 유비퀴놀을 재생성하는데 중요하며, 이는 기생충이 피리미딘 생합성의 특이한 방식을 사용할 수 있도록 한다.
전자전달 플라보단백질:유비퀴논 산화환원효소
전자전달 플라보단백질:유비퀴논 산화환원효소(ETF:Q 산화환원효소) 또는 전자전달 플라보단백질 탈수소효소는 전자전달계의 세 번째 진입 지점이다. ETF:Q 산화환원효소는 미토콘드리아 기질의 전자전달 플라보단백질(electron-transferring flavoprotein, ETF)로부터 전자를 받아서 유비퀴놀을 환원시키는 효소이다. ETF:Q 산화환원효소는 플라빈과 [4Fe–4S] 클러스터를 포함하고 있지만, 다른 호흡 복합체들과는 달리 막의 표면에 부착해 있고, 인지질 이중층을 통과하지 않는다.
-
()

포유류에서 ETF:Q 산화환원효소 대사 경로는 여러 아실-CoA 탈수소효소로부터 전자를 받아들일 수 있기 때문에 지방산의 β 산화와 아미노산과 콜린의 이화작용에서 중요하다. 식물에서 ETF:Q 산화환원효소는 장기간의 어둠에서 생존을 가능하게 하는 대사 반응에서도 중요하다.
유비퀴논:사이토크롬 c 산화환원효소 (복합체 Ⅲ)

유비퀴논:사이토크롬 c 산화환원효소는 사이토크롬 c 환원효소 또는 사이토크롬 bc1 복합체, 또는 간단하게 복합체 III로도 알려져 있다. 포유류에서 복합체 III는 11개의 단백질 소단위체, [2Fe-2S] 철-황 클러스터, 3개의 사이토크롬(1개는 사이토크롬 c1, 2개는 사이토크롬 b)을 포함하는 동일한 소단위체 복합체 2개로 구성된 이합체 효소이다. 사이토크롬은 적어도 하나의 헴기를 갖고 있는 전자전달 단백질의 일종이다. 복합체 III의 헴기 안에 있는 철 원자는 전자가 단백질을 통해 전달됨에 따라 환원된 Fe2+와 산화된 Fe3+ 상태 사이를 번갈아가며 나타내게 된다.
복합체 III 에 의해 촉매되는 반응은 유비퀴놀 1분자의 산화와 미토콘드리아 내막과 느슨하게 결합한 헴단백질인 사이토크롬 c 2분자의 환원이다. 2개의 전자를 운반하는 유비퀴논과는 달리, 사이토크롬 c는 하나의 전자만을 운반한다.
-
()

유비퀴놀(QH2)로부터 사이토크롬 c로 한 번에 하나의 전자가 전달될 수 있기 때문에, 복합체 III의 반응 메커니즘은 다른 호흡 복합체보다 더 정교하며, Q 회로라고 하는 두 단계 반응으로 일어난다. 첫 번째 단계에서 복합체 III는 3가지 기질(유비퀴놀, 사이토크롬 c, 유비퀴논)과 결합하는데 첫 번째 기질인 유비퀴놀(QH2)에서 하나의 전자가 두 번째 기질인 사이토크롬 c로 전달된다. 유비퀴놀(QH2)에서 방출된 2개의 H+(양성자)는 막 사이 공간으로 이동한다. 세 번째 기질은 유비퀴논(Q)이며, 유비퀴논은 유비퀴놀(QH2)로부터 두 번째 전자를 받아들여 세미퀴논 라디칼(Q.−)로 환원된다. 유비퀴놀(QH2)과 유비퀴논(Q)은 방출되지만 세미퀴논 라디칼(Q.−)은 복합체 III에 결합된 채로 남아있는다. 두 번째 단계에서 유비퀴놀(QH2)의 두 번째 분자가 결합되어 다시 첫 번째 전자를 사이토크롬 c로 전달한다. 두 번째 전자는 복합체 III에 결합된 세미퀴논으로 전달되고, 미토콘드리아 기질에서 2개의 H+(양성자)를 얻어서 유비퀴놀(QH2)로 환원되는데 사용된다. 이렇게 생성된 유비퀴놀(QH2)은 복합체 III로부터 방출된다.
유비퀴논이 미토콘드리아 내막의 안쪽 면(N쪽)에서 유비퀴놀로 환원되고, 다른쪽 면(P쪽)에서 유비퀴논으로 산화됨에 따라 미토콘드리아 내막을 가로지르는 H+(양성자)의 순이동이 일어나서 H+(양성자)의 농도 기울기 형성에 기여한다. 복합체 III에서 일어나는 Q 회로의 다소 복잡한 두 단계 메커니즘은 H+(양성자) 운반의 효율성을 증가시키기 때문에 중요하다. Q 회로 대신에 유비퀴놀(QH2) 1분자가 사이토크롬 c 2분자를 직접적으로 환원시키는데 사용된다면, 사이토크롬 c 하나가 환원될 때 단지 하나의 H+(양성자)가 운반되므로 효율성은 절반으로 줄어들게 된다.
사이토크롬 c 산화효소 (복합체 Ⅳ)

사이토크롬 c 산화효소 또는 복합체 IV는 전자전달계의 마지막 단백질 복합체이다. 포유류의 효소는 매우 복잡한 구조를 가지고 있으며, 13개의 소단위체, 2개의 헴기, 구리 원자 3개, 마그네슘 원자 1개, 아연 원자 1개를 포함하는 금속 이온으로 구성된다.
복합체 IV는 전자전달계에서 최종 반응을 수행하며, 미토콘드리아 내막을 가로질러 H+(양성자)를 미토콘드리아 기질에서 막 사이 공간으로 능동수송하면서 전자를 산소로 전달한다. 최종 전자 수용체인 산소(O2)는 이 단계에서 물(H2O)로 환원된다. H+(양성자)를 직접 능동수송하는 것과 산소의 환원과정에서 미토콘드리아 기질의 H+(양성자)를 소비하는 것은 H+(양성자)의 농도 기울기 형성에 기여한다. 촉매되는 반응은 사이토크롬 c의 산화 및 산소의 환원이다.
-
()

대체 산화효소 및 환원효소
많은 진핵생물들은 위에서 설명한 포유류의 효소와 다른 전자전달계를 가지고 있다. 예를 들어, 식물에는 미토콘드리아 기질 쪽이 아닌 막 사이 공간 쪽에서 NADH를 산화시키는 외부 NAD(P)H 탈수소효소가 있으며, 전자를 유비퀴논 풀에 전달한다. 이러한 효소들은 H+(양성자)를 능동수송하지 않으므로 미토콘드리아 내막을 가로지르는 전기화학적 기울기에는 영향을 미치지 않고 유비퀴논을 환원시킨다. 또한 식물의 미토콘드리아 내막에는 기질에 있는 NADH로부터 유비퀴논으로 전자를 직접 전달하는 대체 NADH 탈수소효소도 있다.
전자전달계의 또 다른 예는 식물 뿐만 아니라 일부 균류, 원생생물, 동물에서 발견되는 대체 산화효소이다. 대체 산화효소는 유비퀴놀에서 산소로 전자를 직접 전달한다.
대체 NADH 탈수소효소 및 대체 산화효소에 의한 대체 전자전달 경로는 일반적인 전자전달계에 비해 낮은 ATP 수율을 갖는다. 단축된 대체 전자전달 경로로 인해 얻을 수 있는 이점은 명확하지 않다. 그러나 대체 산화효소는 감기, 활성산소, 병원체에 의한 감염, 전체 전자전달계를 억제하는 다른 인자들과 같은 스트레스에 반응하여 생성된다. 따라서 대체 전자전달 경로는 산화적 스트레스를 줄임으로써 손상에 대한 생물체의 저항력을 향상시킬 수 있다.
복합체들의 구성
호흡 사슬의 복합체들이 어떻게 구성되어 있는지에 대한 최초의 모델은 복합체들이 미토콘드리아 내막에서 자유롭고 독립적으로 확산된다는 것이었다. 그러나 최근의 자료에 따르면 복합체들은 초복합체(supercomplex) 또는 호흡체(respirasome)라고 불리는 고차원적인 구조를 형성하는 것으로 보인다. 이 모델에서 다양한 복합체들은 상호작용하는 효소들의 조직화된 세트로 존재한다. 이러한 연관성은 다양한 효소 복합체들 사이에 기질의 채널링(channeling)을 허용하여 전자전달의 속도와 효율성을 증가시킬 수 있다. 포유류의 초복합체 내의 일부 구성 성분은 다른 구성 성분보다 많은 양으로 존재하며 일부 자료에서는 복합체 I : II : III : IV : ATP 생성효소의 비가 약 1 : 1 : 3 : 7 : 4였다. 그러나 어떤 자료들은 이 모델과 맞지 않는 것처럼 보이기 때문에 이 초복합체 가설에 대한 논쟁은 완전히 해결되지 않았다.
원핵생물의 전자전달계
진핵생물에서 전자전달계의 구조와 기능이 일반적으로 유사성을 가지고 있는 것과는 달리 세균과 고세균은 매우 다양한 전자전달효소들을 가지고 있다. 세균과 고세균은 다양한 화학 물질을 기질로 사용한다. 진핵생물과 마찬가지로 원핵생물의 전자전달계는 기질의 산화로부터 방출된 에너지를 사용하여 막을 가로질러 이온을 능동수송하고 막을 경계로 전기화학적 기울기를 형성시킨다. 세균에서는 대장균(Escherichia coli)의 산화적 인산화가 가장 잘 연구되고 알려져 있는 반면 고세균의 산화적 인산화는 현재까지 잘 알려져 있지 않다.
진핵생물과 원핵생물의 산화적 인산화에서 주된 차이점은 원핵생물은 전자 공여체나 전자 수용체로 매우 다양한 물질을 사용한다는 점이다. 이것은 원핵생물이 다양한 환경 조건하에서 살아갈 수 있게 한다. 예를 들어 대장균에서 산화적 인산화는 아래에 열거된 다양한 환원제와 산화제에 의해 작동할 수 있다. 화학 물질의 표준환원전위는 산화되거나 환원될 때 방출되는 에너지의 양으로 측정한다. 표준환원전위에서 환원제는 음의 값을 가지고, 산화제는 양의 값을 가진다.
| 호흡 효소 | 산화환원 쌍 | 표준환원전위
(V) |
|---|---|---|
| 포름산 탈수소효소 | 탄산수소 이온 / 포름산염 | −0.43 |
| 수소화효소 | 양성자 / 수소 | −0.42 |
| NADH 탈수소효소 | NAD+ / NADH | −0.32 |
| 글리세롤 3-인산 탈수소효소 | 다이하이드록시아세톤 인산 / 글리세롤 3-인산 | −0.19 |
| 피루브산 탈수소효소 | 아세트산 + 이산화 탄소 / 피루브산 | ? |
| 젖산 탈수소효소 | 피루브산 / 젖산 | −0.19 |
| D-아미노산 탈수소효소 | 산소산 + 암모니아 / 아미노산 | ? |
| 퀴노단백질 포도당 탈수소효소 | 글루콘산 / 포도당 | −0.14 |
| 석신산 탈수소효소 | 푸마르산 / 석신산 | +0.03 |
| 유비퀴놀 산화효소 | 산소 / 물 | +0.82 |
| 질산 환원효소 | 질산염 / 아질산염 | +0.42 |
| 아질산 환원효소 | 아질산염 / 암모니아 | +0.36 |
| 다이메틸 설폭사이드 환원효소 | 다이메틸 설폭사이드 / 다이메틸 설파이드 | +0.16 |
| 트라이메틸아민 N-옥사이드 환원효소 | 트라이메틸아민 N-옥사이드 / 트라이메틸아민 | +0.13 |
| 푸마르산 환원효소 | 푸마르산 / 석신산 | +0.03 |
위의 표에서와 같이 대장균은 전자 공여체로 폼산, 수소, 젖산과 같은 환원제를 사용하고 전자 수용체로 질산, 다이메틸 설폭사이드(dimethyl sulfoxide, DMSO), 산소를 이용해 살아갈 수 있다. 산화제와 환원제 간의 표준환원전위 차가 클수록 반응할 때 더 많은 에너지가 방출된다. 이들 화합물들 중에서 석신산/푸마르산 쌍은 표준환원전위 값이 0에 가깝기 때문에 특이하다. 따라서 석신산은 산소와 같은 강한 산화제를 사용하여 푸마르산으로 산화될 수 있고, 푸마르산은 폼산과 같은 강한 환원제를 사용하여 석신산으로 환원될 수 있다. 이러한 대체 반응은 각각 석신산 탈수소효소 및 푸마르산 환원효소에 의해 촉매된다.
일부 원핵생물은 표준환원전위 차가 작은 산화환원 쌍을 사용한다. 예를 들어 니트로박터(Nitrobacter)와 같은 질화세균은 아질산을 질산으로 산화시켜 산소에 전자를 제공한다. 이 반응에서 방출되는 소량의 에너지는 H+(양성자)를 능동수송하고 ATP를 생성하기에 충분하지만, 동화작용에 사용하기 위한 NADH 또는 NADPH를 직접 생산하기에는 충분하지 않다. 이 문제는 아질산 산화환원효소를 사용하여 전자전달계의 일부를 역방향으로 진행시켜 복합체 I이 NADH를 생성하도록 하는 충분한 양성자 구동력을 생성함으로써 해결된다.
원핵생물은 환경 조건에 따라 생성되는 효소를 변화시킴으로써 전자 공여체와 전자 수용체의 사용을 조절한다. 다른 산화효소와 환원효소가 동일한 유비퀴논 풀을 사용하기 때문에 이러한 유연함이 가능하다. 이것은 일반적인 유비퀴놀 중간생성물에 의해 연결된 많은 효소들의 조합이 함께 작용할 수 있게 한다. 따라서 이러한 호흡 사슬은 효소를 쉽게 교체할 수 있는 시스템으로 모듈식 디자인을 갖고 있다.
이러한 대사에서의 다양성에 더하여 원핵생물은 동질효소를 가지고 있다. 예를 들어 대장균에서 산소를 전자 수용체로 사용하는 유비퀴놀 산화효소에는 두 가지 유형이 있다. 고도의 호기성 조건하에서 대장균은 전자당 2개의 H+(양성자)를 운반할 수 있는 산소에 더해 낮은 친화성을 갖는 산화효소를 사용한다. 그러나 산소량이 감소하면 전자당 하나의 H+(양성자)만을 전달하지만, 산소에 대해 높은 친화성을 갖는 산화효소로 전환한다.
ATP 생성효소 (복합체 Ⅴ)
ATP 생성효소 또는 복합체 V는 산화적 인산화 경로의 마지막 효소이다. ATP 생성효소는 거의 모든 생물에서 발견되고 원핵생물과 진핵생물에서 동일한 방식으로 작동한다. ATP 생성효소는 ADP와 인산(Pi)으로부터 ATP를 합성하기 위해 막을 경계로 형성된 H+의 농도 기울기의 형태로 저장된 에너지를 사용한다. 하나의 ATP를 합성하는데 필요한 H+(양성자)의 수는 3~4 사이로 추정되며, 일부 세포들은 다른 조건들에 맞추어 이 비율을 바꿀 수 있다.
-
()

이 인산화 반응은 양성자 구동력이 변화됨에 따라 평형이 이동될 수 있다. 양성자 구동력이 없으면 ATP를 가수분해해서 H+(양성자)를 미토콘드리아 기질에서 막 사이 공간으로 막을 가로질러 능동수송하고, ATP 생성효소 반응은 오른쪽에서 왼쪽으로 진행된다. 그러나 양성자 구동력이 높을 때 H+(양성자)가 농도 기울기에 따라 ATP 생성효소를 통해 확산되어 나갈 때 ADP가 ATP로 전환되고, 반응은 왼쪽에서 오른쪽으로 진행된다. 식물의 액포에 존재하는 V형 ATPase는 ATP를 가수분해하면서 H+(양성자)를 능동수송해서 액포 내부를 산성화시킨다.
ATP 생성효소는 버섯 모양의 거대한 단백질 복합체이다. 포유류의 ATP 생성효소는 16개의 소단위체를 포함하고, 약 600 kDa의 분자량을 갖는다. Fo(o는 올리고마이신 민감성을 의미)는 막에 묻혀있는 부분으로 c 소단위체와 양성자(H+) 통로를 포함한다. F1은 기둥과 공 모양의 머리 부분으로 ATP를 합성하는 부위이다. F1은 9개의 소단위체로 구성되어 있고, 소단위체는 5종류가 있으며, 그 구성은 α3β3γδε이다. F1의 공 모양의 복합체는 2가지 종류의 6개의 단백질(3개의 α 소단위체와 3개의 β 소단위체)로 구성된다. F1의 γ 소단위체는 α 소단위체와 β 소단위체로 구성된 공 모양의 구조물 중앙을 관통하는 중심축 같은 구조이다. F1의 α 및 β 소단위체는 둘 다 뉴클레오타이드와 결합하지만, β 소단위체만이 ATP 합성반응을 촉매한다. F1의 γ 소단위체와 ε 소단위체는 α 소단위체와 β 소단위체를 효소의 기저부에 고정시킨다.
H+(양성자)가 ATP 생성효소의 기저부에 있는 통로를 통해 막을 가로질러 통과함에 따라 Fo의 양성자 구동 모터가 회전한다. 모터의 회전은 H+(양성자)가 통로를 지나갈 때 c 소단위체의 고리를 회전시키는 정전기적 상호작용과 이로 인한 c 소단위체 고리 내에서 아미노산의 이온화 상태의 변화로 인해 야기될 수 있다. 이러한 회전 고리는 α 소단위체와 β 소단위체를 관통하는 중심축(γ 소단위체)을 차례로 회전시킨다. α 와 β 소단위체는 고정자 역할을 하는 측면 팔 구조에 의해 스스로 회전하는 것을 방지한다. α 와 β 소단위체를 관통하는 γ 소단위체의 움직임은 β 소단위체 내의 활성 부위에서 ATP를 생성하고 ATP를 방출하기 위해 필요한 에너지를 제공한다.

이러한 ATP 합성반응은 결합 변화 메커니즘이라고 불리며 각각의 β 소단위체가 3가지 입체 형태를 순환하도록 유도한다. "열린" 상태에서 ADP와 인산(Pi)은 활성 부위로 들어간다(그림에서 진한 붉은색으로 표시됨). "느슨한" 상태에서 단백질은 분자들과 느슨하게 결합한다(주황색으로 표시). 효소는 다시 구조를 변화시키고 새로 생성된 ATP를 효소의 활성부위에 매우 높은 친화력으로 결합시킨다("단단한" 상태, 분홍색으로 표시). 마지막으로 효소의 활성 부위에서 ATP가 방출되고, 다시 "열린" 상태로 되돌아가 다음 사이클을 반복한다.
일부 세균과 고균에서 ATP 합성은 H+(양성자)의 이동이 아니라 나트륨 이온(Na+)의 막을 통한 이동에 의해서 이루어진다.Methanococcus와 같은 고균세은 다른 세균 및 진핵생물의 ATP 생성효소 소단위체와는 다른 단백질을 포함하는 A1Ao 생성효소를 갖고 있다. 일부 종에서 A1Ao 형태의 효소가 나트륨(Na+)으로 구동되는 ATP 생성효소일 수도 있지만, 이는 모든 경우에서 다 그러한 것은 아니다.
활성산소
산소(O2)는 강력한 산화제이기 때문에 이상적인 최종 전자 수용체이다. 산소의 환원 과정은 잠재적으로 해로운 중간생성물의 형성을 포함한다. 4개의 전자와 4개의 양성자가 산소(O2)로 전달되면 물(H2O)로 환원되는데, 이는 무해하다. 하지만 1개 또는 2개의 전자가 전달되면 초과산화물 또는 과산화물을 생성하게 되는데, 이들은 반응성이 커서 위험하다.
-
()
![{\displaystyle {\begin{matrix}{}\\{\ce {O2->[{\ce {e^{-}}}]{\underset {Superoxide}{O2^{\underline {\bullet }}}}->[{\ce {e^{-}}}]{\underset {Peroxide}{O2^{2-}}}}}\\{}\end{matrix}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/d22cb3bd5583f49decea45c79baedb470f5b2fe3)
이러한 활성산소들과 하이드록시 라디칼과 같은 반응 산물들은 단백질을 산화시키고 DNA에 돌연변이를 유발하기 때문에 세포에 매우 해롭다. 이러한 세포 손상은 질병의 원인이 될 수 있으며, 노화의 원인들 중 하나로 제시된다.
사이토크롬 c 산화효소 복합체는 산소(O2)를 물(H2O)로 환원시키는데 매우 효율적이며, 부분적으로 환원된 중간생성물을 거의 방출하지 않는다. 그러나 전자전달계에 의해 소량의 초과산화물 및 과산화물이 생성된다. 반응성이 큰 세미퀴논 라디칼이 Q 회로에서 중간생성물로 형성되기 때문에 복합체 III에서 유비퀴논의 환원이 특히 중요하다. 세미퀴논 라디칼은 불안정해서 전자가 산소로 직접 전달될 때 전자의 누출을 일으켜서 초과산화물을 형성시킨다. 이러한 양성자 펌핑 복합체에 의한 활성산소의 생성은 높은 막전위에서 가장 크기 때문에 미토콘드리아가 ATP 생산과 산화제 생성의 균형을 이루는 좁은 범위 내에서 막전위를 유지하도록 이들의 활성을 조절하는 것이 제안되었다. 예를 들어 산화제는 막전위를 감소시키는 짝풀림 단백질을 활성화시킬 수 있다.
이러한 활성산소에 대응하기 위해 세포에는 비타민 C, 비타민 E와 같은 항산화 비타민과 초과산화물 불균등화효소, 카탈레이스, 과산화효소와 같은 항산화효소를 포함하는 수 많은 항산화 시스템을 가지고 있어서 활성산소를 해독하여 세포 손상을 방지한다.
저해제
산화적 인산화를 저해하는 잘 알려진 몇 가지 약물과 독소가 있다. 이들 독소 중 어느 하나가 산화적 인산화 경로에서 하나의 효소만을 저해하더라도 이러한 임의의 단계의 저해는 다른 나머지 과정들까지 정지시킨다. 예를 들어 올리고마이신이 ATP 생성효소를 저해하면 H+(양성자)는 미토콘드리아 기질로 되돌아갈 수 없다. 결과적으로 H+의 농도 기울기가 너무 강해져서 펌프가 이를 극복할 수 없기 때문에, 양성자 펌프가 작동할 수 없게 된다. NADH가 더 이상 산화되지 못하고 NAD+의 농도가 효소가 사용할 수 있는 농도 이하로 떨어지게 되면 시트르산 회로의 작동이 중단된다.
| 화합물 | 용도 | 작용 부위 | 산화적 인산화에 미치는 영향 |
|---|---|---|---|
|
사이안화물 일산화 탄소 아자이드화물 황화 수소 |
독 | 복합체 IV | 사이토크롬 c 산화효소의 Fe-Cu 중심에 산소보다 더 강하게 결합하여 전자전달계를 저해하여 산소의 환원을 방해한다. |
| 올리고마이신 | 항생제 | 복합체 V | Fo을 통한 H+(양성자)의 흐름을 차단하여 ATP 생성효소를 저해한다. |
|
CCCP 2,4-다이나이트로페놀 |
독, 체중 감소 | 미토콘드리아 내막 | 이오노포어는 막을 가로질러 H+(양성자)를 운반함으로써 H+(양성자) 농도 기울기 형성을 방해한다. 이오노포어는 H+(양성자)의 펌핑과 ATP 합성을 짝풀리게 한다. |
| 로테논 | 농약 | 복합체 I | 유비퀴논 결합 부위를 차단하여 복합체 I에서 유비퀴논으로의 전자전달을 방해한다. |
| 말론산과 옥살아세트산 | 독 | 복합체 II | 석신산 탈수소효소(복합체 II)의 경쟁적 저해제. |
| 안티마이신 A | 살어제 | 복합체 III | 사이토크롬 c 환원효소에 결합하여 유비퀴놀의 산화를 저해한다. |
산화적 인산화 과정의 모든 저해제가 독소는 아니다. 갈색지방조직에서 짝풀림 단백질이라고 불리는 조절된 H+(양성자) 통로는 호흡을 ATP 합성으로부터 분리할 수 있다. 이러한 짝풀림 호흡은 열을 생산하며, 특히 동면동물에서 체온을 유지하는 방법으로 중요하다.
역사
산화적 인산화에 대한 연구는 발효에서 인산이 중요한 역할을 한다는 아서 하든의 1906년 논문에서 시작되었지만, 처음에는 당인산만이 관련되어 있는 것으로 알려졌었다. 그러나 1940년대 초에 헤르만 칼카르(Herman Kalckar)에 의해 당의 산화와 ATP의 생성 사이의 연관성이 확립되었고, 1941년 프리츠 앨버트 리프만이 에너지 전달 과정에서 ATP의 중심적인 역할을 확인했다. 이후 1949년에 모리스 프리드킨(Morris Friedkin)과 앨버트 L. 레닌저(Albert L. Lehninger)는 조효소인 NADH가 시트르산 회로 및 ATP 합성과 같은 대사 경로와 연결되어 있다는 것을 증명했다. 산화적 인산화라는 용어는 1939년 소련의 볼로디미르 벨리체르(Volodymyr Belitser)가 만들었다.
이 후 20년 동안 ATP가 생성되는 메커니즘은 수수께끼로 남아있었고, 과학자들은 산화와 인산화 반응을 연결할 수 있는 "고에너지 중간산물"을 찾으려고 노력했다. 이러한 수수께끼는 1961년 피터 D. 미첼이 화학삼투설을 발표함으로써 해결되었다. 처음에 화학삼투설은 과학자들 사이에서 논란이 많았지만 서서히 받아들여졌고, 미첼은 1978년에 노벨 화학상을 수상했다. 후속 연구들은 ATP 생성효소에 대한 에프라임 랙커(Efraim Racker)의 연구, 전자전달계의 복합체에 대한 데이비드 E. 그린(David E. Green)의 연구처럼 관련 효소들을 정제하고 특징짓는 데에 집중되었다.폴 D. 보이어는 1973년에 결합 변화 메커니즘을, 1982년에 회전 촉매 방식을 제안하여 ATP 생성효소의 작동 메커니즘을 해결하기 위한 이론들을 제시했다. 이 후에 존 E. 워커는 산화적 인산화에 관여하는 효소들의 구조를 연구했다. 존 E. 워커와 폴 D. 보이어는 1997년에 노벨 화학상을 수상했다.
같이 보기
주해
더 읽을거리
입문 서적
- Nelson DL; Cox MM (2004). 《Lehninger Principles of Biochemistry》 4판. W. H. Freeman. ISBN 0-7167-4339-6.
- Schneider ED; Sagan D (2006). 《Into the Cool: Energy Flow, Thermodynamics and Life》 1판. University of Chicago Press. ISBN 0-226-73937-6.
- Lane N (2006). 《Power, Sex, Suicide: Mitochondria and the Meaning of Life》 1판. Oxford University Press, USA. ISBN 0-19-920564-7.
고급 서적
- Nicholls DG; Ferguson SJ (2002). 《Bioenergetics 3》 1판. Academic Press. ISBN 0-12-518121-3.
- Haynie D (2001). 《Biological Thermodynamics》 1판. Cambridge University Press. ISBN 0-521-79549-4.
- Rajan SS (2003). 《Introduction to Bioenergetics》 1판. Anmol. ISBN 81-261-1364-2.
- Wikstrom M (Ed) (2005). 《Biophysical and Structural Aspects of Bioenergetics》 1판. Royal Society of Chemistry. ISBN 0-85404-346-2.
일반적인 내용
- Animated diagrams illustrating oxidative phosphorylation Wiley and Co Concepts in Biochemistry
- On-line biophysics lectures Antony Crofts, University of Illinois at Urbana-Champaign
- ATP Synthase Graham Johnson
구조에 관한 내용
-
PDB molecule of the month:
- ATP synthase Archived 2020년 7월 24일 - 웨이백 머신
- Cytochrome c Archived 2020년 7월 24일 - 웨이백 머신
- Cytochrome c oxidase Archived 2020년 7월 24일 - 웨이백 머신
- Interactive molecular models at Universidade Fernando Pessoa: