
전자전달계(電子傳達系, 영어: electron transport chain, ETC)는 산화환원반응(산화와 환원이 동시에 일어남)을 통해 전자를 전자공여체로부터 전자수용체로 전달하고 이러한 전자전달을 막을 가로질러 양성자(H+ 이온)의 능동수송과 짝짓게 만드는 일련의 단백질 복합체들이다. 일련의 단백질들은 미토콘드리아 내막에 존재한다. 전자는 NADH와 FADH2로부터 4개의 다중 소단위체로 구성된 대형 효소 복합체와 2개의 이동 가능한 전자 운반체를 포함하고 있는 전자전달계로 전달된다. 전자전달계에 존재하는 많은 효소들는 막에 결합되어 있다.
전자전달계를 통한 전자의 흐름은 에너지 방출반응이다. 산화환원반응의 에너지는 아데노신 삼인산(ATP)의 합성을 유도하는 양성자의 전기화학적 기울기를 형성한다. 산소 호흡에서 전자전달의 최종 전자수용체는 산소(O2)이다. 무산소 호흡에서는 황산염과 같은 다른 전자 수용체가 사용된다.
전자전달계에서 산화환원반응은 반응물과 생성물의 깁스 자유 에너지의 차이에 의해 일어난다. 더 높은 에너지 상태의 전자공여체와 전자수용체가 더 낮은 에너지 상태의 생성물로 전환될 때 방출되는 자유 에너지는 전자가 더 낮은 산화환원전위에서 더 높은 산화환원전위로 이동하는 동안 전자전달계의 복합체들에 의해 H+ 이온의 전기화학적 기울기를 생성하는 데 사용된다. 이렇게 전자전달과 화학삼투에 의한 ATP의 생성을 짝지어서, 막을 경계로 형성된 H+ 이온의 농도 기울기를 이용하여 ATP를 생성한다.
진핵생물에서 전자전달계와 ATP 생성효소는 미토콘드리아 내막에서 발견된다. 세포 호흡에서 환원된 전자 운반체(예: NADH 및 FADH2)에 저장된 에너지는 전자전달계에 의해 양성자(H+)를 막 사이 공간으로 능동수송하며 미토콘드리아 내막을 경계로 전기화학적 기울기를 형성하는 데 사용된다. 광합성 진핵생물에서 전자전달계는 틸라코이드 막에서 발견된다. 여기서 빛 에너지는 물로부터 공급받는 전자를 고에너지 전자로 만들어 전자전달계로 전달하고, 전자전달 과정에서 형성된 H+의 농도 기울기를 이용해 ATP를 생성한다. 세균에서 전자전달계는 종에 따라 다를 수 있지만, 전자전달을 통한 전기화학적 기울기 형성 및 ATP 생성효소를 통한 화학삼투와 ATP 합성을 짝을 짓는 산화적 인산화와 일련의 산화환원반응들은 공통적이다.
미토콘드리아의 전자전달계
대부분의 진핵세포는 시트르산 회로, 지방산의 β 산화, 아미노산의 산화의 생성물로부터 ATP를 합성하는 미토콘드리아를 가지고 있다. 미토콘드리아 내막에서 NADH와 FADH2의 고에너지 전자는 전자전달계를 통해 최종 전자 수용체인 산소(O2)로 전달되어 물(H2O)로 환원된다. 전자전달계는 일련의 효소적 전자공여체 및 전자수용체들로 구성된다. 각 전자공여체는 전자를 보다 전기음성도가 큰 전자수용체로 전달하고, 이 과정은 전자가 전자전달계에서 전기음성도가 가장 큰 최종 전자수용체인 산소(O2)로 전달될 때까지 계속 진행된다. 전자공여체와 전자수용체 사이의 전자전달 과정에서 단계적으로 에너지를 방출하게 되는데, 이 에너지는 미토콘드리아 내막을 가로질러 양성자(H+)를 미토콘드리아 기질에서 막 사이 공간으로 능동수송하여 양성자 농도 기울기를 형성하는 데 사용되어 일을 할 수 있는 열역학적 상태를 만든다. 전자전달계의 산화환원반응에 의해 확립된 전기화학적 기울기를 사용하여 ADP를 ATP로 인산화시키기 때문에 이러한 전체적인 과정을 산화적 인산화라고 부른다.
미토콘드리아의 산화환원 운반체
전자전달계에서 전자전달을 통해 얻은 에너지는 미토콘드리아 기질에서 막 사이 공간으로 양성자(H+)를 능동수송하여 미토콘드리아 내막을 경계로 전기화학적 양성자 농도 기울기를 생성하는 데 사용된다. 전기화학적 기울기로 형성된 양성자 구동력은 H+의 화학적 농도 차이(ΔpH)와 막을 경계로 형성된 전기적 전위차(ΔΨ)로 구성된다. 그러나 미토콘드리아에서의 H+ 농도 기울기는 미토콘드리아 막 전위(ΔΨM)에는 크게 영향을 미치지 않는다. 전기화학적 기울기로 형성된 전기화학적 위치 에너지는 H+가 ATP 생성효소를 통해 막 사이 공간(높은 H+ 농도)에서 미토콘드리아 기질(낮은 H+ 농도)로 확산되는 흐름(화학삼투)을 형성하여 ADP와 무기 인산(Pi)로부터 ATP를 생성하도록 한다. 복합체 I(NADH:유비퀴논 산화환원효소)은 고에너지 전자 운반체인 NADH로부터 전자를 받아 유비퀴논(조효소 Q)으로 전달한다. 유비퀴논은 복합체 II(석신산 탈수소효소)로부터도 전자를 전달받는다. 유비퀴논은 전자를 복합체 III(사이토크롬 bc1 복합체)로 전달하고, 복합체 III는 전자를 사이토크롬 c로 전달한다. 사이토크롬 c는 전자를 복합체 IV(사이토크롬 c 산화효소)로 전달하며, 복합체 IV는 전자와 수소 이온(H+)을 사용하여 산소 분자(O2)를 물(H2O)로 환원시킨다.
미토콘드리아에서 4개의 막 결합 복합체가 확인되었다. 각각의 복합체는 미토콘드리아 내막에 파묻혀 있는 매우 복잡한 막관통 구조를 가지고 있다. 이 중 복합체 I, III, IV는 양성자 펌프이다. 구조는 지용성 전자 운반체와 수용성 전자 운반체에 의해 전기적으로 연결된다. 전자전달계의 전체적인 개요는 다음과 같다.
NADH+H+ → 복합체 I → 유비퀴논(Q)
↑
복합체 II
↑
석신산 → 복합체 III → 사이토크롬 c → 복합체 IV → H2O
↑
복합체 II
↑
석신산
복합체 I
복합체 I(NADH:유비퀴논 산화환원효소, I형 NADH 탈수소효소, 미토콘드리아 복합체 I) (EC 1.6.5.3)에서 2개의 전자가 NADH로부터 제거되어 지용성 전자 운반체인 유비퀴논(Q)으로 전달된다. 환원된 생성물인 유비퀴놀(QH2)은 막 내에서 자유롭게 확산되고 복합체 I은 4개의 양성자(H+)를 능동수송하여 양성자의 농도 기울기를 형성시킨다. 복합체 I은 산소로의 조기 전자 누출이 일어나는 주요 부위 중 하나이며, 따라서 초과산화물 생성의 주요 부위 중 하나이다.
전자전달의 경로는 다음과 같다.
NADH는 2개의 전자로 플라빈 모노뉴클레오타이드(FMN)를 FMNH2로 환원시킴으로써 니코틴아마이드 아데닌 다이뉴클레오타이드(NAD+)로 산화된다. FMNH2는 1개의 전자를 잃고 세미퀴논 중간생성물 상태로 산화되며, 또 다른 1개의 전자를 잃어 FMN으로 산화된다. 따라서 각각의 전자는 FMNH2로부터 Fe-S 클러스터로, Fe-S 클러스터로부터 유비퀴논(Q)로 전달된다. 첫 번째 전자전달은 유비퀴논(Q)의 자유 라디칼(세미퀴논) 형태를 생성하고, 두 번째 전자전달은 세미퀴논 형태를 유비퀴놀 형태인 QH2로 환원시킨다. 이 과정에서 4개의 양성자(H+)가 미토콘드리아 기질로부터 막 사이 공간으로 능동수송된다. 전자가 복합체 전체에서 지속적으로 산화되고 환원됨에 따라 전자의 흐름이 막 내 복합체의 180 Å의 폭을 따라 생성된다. 이러한 전자의 흐름은 NADH로부터 전자 2개당 4개의 양성자(H+)를 막 사이 공간으로 능동수송시킨다.
복합체 II
복합체 II(석신산 탈수소효소, 석신산-CoQ 환원효소) (EC 1.3.5.1)에서 전달되는 전자는 석신산에서 시작된 퀴논 풀(Q)로 전달되어 플라빈 아데닌 다이뉴클레오타이드(FAD)를 통해 Q로 전달된다. 복합체 II는 석신산 탈수소효소(SDHA), 미토콘드리아의 석신산 탈수소효소 [유비퀴논] 철-황 소단위체(SDHB), 석신산 탈수소효소 복합체 소단위체 C(SDHC), 석신산 탈수소효소 복합체 소단위체 D(SDHD)의 4개의 단백질 소단위체로 구성된다. 다른 전자공여체(예: 지방산, 글리세롤 3-인산)도 전자를 FAD를 통해 Q로 전달한다. 복합체 II와 복합체 I은 서로 병렬적인 전자전달 경로를 형성하지만, 복합체 I과는 달리 복합체 II는 막 사이 공간으로 양성자(H+)를 능동수송하지 않는다. 따라서 복합체 II를 통하는 경로는 전체적인 전자전달계의 과정에 기여하는 에너지가 복합체 I을 통하는 경로보다 적다.
복합체 III
복합체 III(사이토크롬 bc1 복합체, CoQH2-사이토크롬 c 환원효소) (EC 1.10.2.2)에서 Q 회로는 양성자의 비대칭적인 흡수/방출에 의해 양성자의 농도 기울기 형성에 기여한다. 2개의 전자는 QO 부위의 QH2로부터 제거되고 막 사이 공간 내에 위치한 수용성 전자 운반체인 두 개의 사이토크롬 c 분자로 순차적으로 전달된다. 다른 2개의 전자는 단백질을 가로질러 유비퀴논의 퀴논 부분이 퀴놀로 환원되는 Qi 부위로 순차적으로 전달된다. 양성자의 농도 기울기는 Qo 부위에서 1개의 퀴놀()의 산화에 의해 Qi 부위에서 1개의 퀴논()이 생성되면서 형성된다. 총 4개의 양성자가 전위되는 데 2개의 양성자는 퀴논을 퀴놀로 환원시키는 데 사용되고 2개의 양성자는 2개의 유비퀴놀 분자로부터 방출된다.








전자전달이 감소하면(높은 막 전위 또는 안티마이신 A와 같은 세포 호흡 저해제에 의해) 복합체 III는 전자를 산소 분자(O2)로 누출시켜 초과산화물을 형성할 수 있다.
복합체 III는 다이머카프롤(BAL, British Antilewisite), 나프토퀴논, 안티마이신에 의해 저해된다.
복합체 IV
복합체 IV(사이토크롬 c 산화효소, 때때로 사이토크롬 AA3라고도 부름) (EC 1.9.3.1)에서 사이토크롬 c의 4개의 분자로부터 제거된 4개의 전자가 산소 분자(O2)로 전달되어 2개의 물(H2O) 분자가 생성된다. 복합체 IV는 배위된 구리 이온과 여러 헴기를 포함하고 있다. 동시에 8개의 양성자(H+)가 미토콘드리아 기질로부터 제거되어(4개만 막을 가로질러 능동수송됨) 양성자의 농도 기울기 형성에 기여한다. 복합체 IV의 양성자 펌핑에 대한 정확한 세부 사항은 여전히 연구 중에 있다. 사이안화물은 복합체 IV의 저해제이다.
전자전달과 화학삼투에 의한 ATP 합성의 짝지음
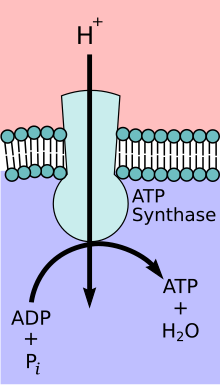
노벨 화학상 수상자인 피터 D. 미첼이 제안한 화학삼투적 인산화 이론은 전자전달계와 화학삼투에 의한 ATP의 합성이 미토콘드리아 내막을 가로질러 형성된 양성자의 농도 기울기에 의해 짝지워져 있다는 것이다. 미토콘드리아 기질에서 막 사이 공간으로 양성자(H+)가 능동수송되어 미토콘드리아 내막을 경계로 전기화학적 기울기(양성자의 농도 기울기)가 형성된다. 이러한 양성자의 농도 기울기에 의해 양성자(H+)가 ATP 생성효소를 통해 막 사이 공간에서 미토콘드리아 기질로 확산(화학삼투)될 때 미토콘드리아 기질 쪽에서 ATP가 합성된다. ATP 생성효소는 때때로 전자전달계의 복합체 V로 설명되기도 한다. ATP 생성효소의 FO 성분은 미토콘드리아 기질로 양성자를 확산시키는 이온 통로 역할을 한다. FO 성분은 a, b, c 소단위체로 구성된다. 미토콘드리아의 막 사이 공간의 양성자는 먼저 소단위체 통로를 통해 ATP 생성효소 복합체로 들어간다. 그런 다음 양성자는 c 소단위체로 이동한다. ATP 생성효소가 가지고 있는 c 소단위체의 갯수는 FO가 1회전을 하도록 만드는 데 필요한 양성자의 갯수를 결정한다. 예를 들어 사람의 ATP 생성효소는 8개의 c 소단위체가 있으므로 FO을 1회전 시키는 데 8개의 양성자(H+)가 필요하다. c 소단위체를 지나 양성자는 마침내 미토콘드리아 기질로 연결된 소단위체 통로를 사용하여 미토콘드리아 기질로 들어간다. 이러한 양성자의 환류는 산화된 형태의 전자 운반체(NAD+ 및 Q)가 생성되는 동안 생성된 자유 에너지를 방출한다. 자유 에너지는 ATP 생성효소 복합체의 F1 성분에 의해 촉매되는 ATP 합성에 사용된다.
전자전달(산화적)과 화학삼투에 의한 ATP 합성(인산화)의 짝지어짐은 ATP 생성의 핵심적인 단계이다. 그러나 특정 경우에는 두 과정의 짝을 풀리게 하는 것이 생물학적으로 유용할 때가 있다. 갈색지방조직의 미토콘드리아 내막에 존재하는 짝풀림 단백질인 써모제닌은 미토콘드리아 기질로 양성자를 흐르게 하는 대체 경로를 제공한다. 또한 티록신은 천연의 짝풀림제이다. ATP 생성효소를 통하지 않는 이러한 양성자의 대체 흐름은 ATP 생성이 아닌 열을 발생시킨다.
역전자 흐름
역전자 흐름은 역산화환원반응에 의한 전자전달계를 통한 전자의 이동이다. 일반적으로 상당한 양의 에너지를 사용해야 하므로 산화된 형태의 전자공여체를 환원시킬 수 있다. 예를 들어 NAD+는 복합체 I에 의해 NADH로 환원될 수 있다. 역전자 흐름을 유도하는 것으로 밝혀진 몇 가지 요인들이 있다. 그러나 이를 확인하려면 더 많은 작업이 필요하다. 그러한 예 중 하나는 ATP 생성효소에 의한 ATP 생성을 차단함으로써 양성자를 축적시켜서 양성자 구동력을 높이게 되고 역전자 흐름을 유도한다.
세균의 전자전달계
진핵생물에서 NADH는 가장 중요한 전자공여체이다. 전자전달계는 "NADH → 복합체 I → Q → 복합체 III → 사이토크롬 c → 복합체 IV → O2"의 순서이며, 여기서 복합체 I, 복합체 III, 복합체 IV는 양성자 펌프이고, 유비퀴논(Q) 및 사이토크롬 c는 이동성 전자운반체이다. 최종 전자수용체는 산소 분자(O2)이다.
원핵생물(세균 및 고세균)에서는 여러 전자공여체와 여러 전자수용체가 있기 때문에 상황이 더 복잡하다. 세균의 일반적인 전자전달계는 다음과 같다.
전자공여체 전자공여체 전자공여체
↓ ↓ ↓
탈수소효소 → 퀴논 → bc1 → 사이토크롬
↓ ↓
산화효소(환원효소) 산화효소(환원효소)
↓ ↓
전자수용체 전자수용체
전자는 탈수소효소 단계, 퀴논 풀의 단계, 이동성 사이토크롬 전자 운반체의 단계의 3가지 단계에서 전자전달계로 들어갈 수 있다. 이들 단계는 연속적으로 더 큰 양의 산화환원전위에 해당하거나 최종 전자수용체에 비해 연속적으로 감소된 전위차에 해당한다. 즉, 전체 산화환원반응의 공여체 → 수용체에 대해 연속적으로 더 작은 깁스 자유 에너지 변화에 해당한다.
세균은 종종 동시에 여러 전자전달계를 사용한다. 세균은 여러 가지 다른 전자공여체, 여러 가지 다른 탈수소효소, 여러 가지 다른 산화효소와 환원효소, 여러 가지 다른 전자수용체를 사용할 수 있다. 예를 들어 대장균(산소가 충분한 조건에서 포도당을 에너지원으로 하여 생장할 때)은 2개의 다른 NADH 탈수소효소와 2개의 다른 퀴놀 산화효소를 사용하여 총 4개의 다른 전자전달계를 동시에 작동시킨다.
모든 전자전달계의 공통적인 특징은 막에 전기화학적 기울기를 생성하는 양성자 펌프가 존재한다는 점이다. 세균의 전자전달계는 미토콘드리아와 같이 3개의 양성자 펌프를 포함할 수도 있고, 1개 또는 2개만 포함할 수도 있다. 세균의 전자전달계는 적어도 1개의 양성자 펌프를 포함하고 있다.
전자공여체
오늘날 생물권에서 가장 일반적인 전자공여체는 유기 분자이다. 유기 분자를 전자 공급원으로 사용하는 생물을 유기영양생물이라고 한다. 유기영양생물(동물, 균류, 원생생물)과 광영양생물(식물과 조류)는 모든 친숙한 생명체들의 대부분을 구성한다.
일부 원핵생물은 무기물을 에너지원으로 사용할 수 있다. 이러한 생물을 무기영양생물이라고 한다. 무기 전자공여체로는 수소, 일산화 탄소, 암모니아, 아질산염, 황, 황화물, 산화 망가니즈, 철이 있다. 무기영양생물은 지구 표면의 수천 미터 아래의 암석에서 서식하는 것으로 발견되었다. 분포 양으로 보면 무기영양생물은 생물권의 유기영양생물과 광영양생물보다 많을 수도 있다.
에너지원으로 무기 전자공여체를 사용하는 것은 진화 연구에서 특히 중요하다. 이러한 유형의 물질대사는 논리적으로 추론해 봤을 때 유기 분자를 에너지원으로 사용하기 전에 존재했음이 틀림없다.
복합체 I과 복합체 II
세균은 다양한 전자공여체를 사용할 수 있다. 유기물이 에너지원인 경우 전자공여체는 NADH 또는 석신산일 수 있으며, 이 경우 전자는 NADH 탈수소효소(미토콘드리아의 복합체 I과 유사) 또는 석신산 탈수소효소(미토콘드리아의 복합체 II와 유사)를 통해 전자전달계로 들어간다. 폼산 탈수소효소, 젖산 탈수소효소, 글리세르알데하이드 3-인산 탈수소효소, H2 탈수소효소(수소화효소)와 같이 다른 에너지원을 처리하기 위해 다른 탈수소효소를 사용할 수 있다. 일부 탈수소효소도 또한 양성자 펌프이다. 다른 탈수소효소는 퀴논 풀로 전자를 이동시킨다. 대부분의 탈수소효소는 세포가 생장하는 환경에 의해 유발되는 대사 요구에 반응하여 세균 세포에서 발현이 유도된다. 대장균의 젖산 탈수소효소의 경우 효소는 호기적으로 사용되며, 다른 탈수소효소와 함께 사용된다. 이는 유도 가능하며 세포에 고농도의 젖산이 존재할 때 발현된다.
퀴논 운반체
퀴논은 막에 존재하는 거대하고 상대적으로 움직이지 않는 거대분자 복합체 사이에서 전자(및 양성자)를 운반하는 이동성 지용성 운반체이다. 세균은 유비퀴논(조효소 Q, 미토콘드리아가 사용하는 것과 동일한 퀴논)과 메나퀴논(비타민 K2)와 같은 관련된 퀴논들을 사용한다. 술폴로부스속의 고세균은 칼다리엘라퀴논을 사용한다. 다른 퀴논을 사용하는 것은 약간 변형된 산화환원전위 때문이다. 산화환원전위의 이러한 변화는 퀴논 구조의 변화로 인해 발생한다. 이들 퀴논의 산화환원전위의 변화는 전자수용체의 변화 또는 세균의 복합체의 산화환원전위의 변화에 적합할 수 있다.
양성자 펌프
양성자 펌프는 막을 경계로 양성자의 농도 기울기를 생성하도록 하는 막 단백질이다. 양성자(H+)는 물리적으로 막을 가로질러 이동할 수 있다. 이것은 미토콘드리아 복합체 I, 복합체 III, 복합체 IV에서 볼 수 있다. 전자를 반대 방향으로 이동시켜도 동일한 효과를 얻을 수 있다. 그 결과 세포질에서 양성자가 감소하고 주변 세포질에서 양성자가 증가한다. 미토콘드리아의 복합체 III는 퀴논에 의해 매개되는(Q 회로) 두 번째 유형의 양성자 펌프를 사용한다.
일부 탈수소효소는 양성자 펌프이나 다른 것들은 그렇지 않다. 대부분의 산화효소와 환원효소는 양성자 펌프이지만, 일부는 그렇지 않다. 세균의 사이토크롬 bc1 복합체는 전부는 아니지만 많은 세균에서 발견되는(대장균에서는 발견되지 않음) 양성자 펌프이다. 이름에서 알 수 있듯이 세균의 사이토크롬 bc1 복합체는 미토콘드리아의 사이토크롬 bc1 복합체(복합체 III)와 유사하다.
사이토크롬 전자 운반체
사이토크롬은 철을 함유하고 있는 단백질이다. 사이토크롬은 두 가지의 매우 다른 환경에서 발견된다.
일부 사이토크롬은 막에 내재되어 있는 크고 이동성이 없는 거대분자 구조의 수용성 운반체로 전자를 왕복수송한다. 미토콘드리아의 이동성 사이토크롬 전자 운반체는 사이토크롬 c이다. 세균은 다양한 이동성 사이토크롬 전자 운반체를 사용한다.
다른 사이토크롬들은 복합체 III와 복합체 IV와 같은 거대분자 내에서 발견된다. 또한 이들은 전자 운반체로 기능하지만, 매우 다른 분자 내 고체 상태의 환경에서도 작동한다.
전자는 이동성 사이토크롬 또는 퀴논 운반체 수준에서 전자전달계로 들어갈 수 있다. 예를 들어 무기 전자공여체(아질산염, 철)의 전자는 사이토크롬 수준에서 전자전달계로 들어간다. 전자가 NADH보다 더 큰 산화환원 수준으로 들어가면 전자전달계가 역으로 작동하여 필요한 고에너지 분자를 생성한다.
말단 산화효소 및 환원효소
세균이 호기성 환경에서 생장할 때 최종 전자수용체인 산소(O2)는 산화효소에 의해 물(H2O)로 환원된다. 세균이 혐기성 환경에서 생장할 때 최종 전자수용체는 환원효소에 의해 환원된다. 미토콘드리아에서 최종 막 복합체(복합체 IV)는 사이토크롬 산화효소이다. 호기성 세균은 다양한 말단 산화효소를 사용한다. 예를 들어 대장균(통성혐기성 생물)에는 사이토크롬 산화효소나 사이토크롬 bc1 복합체가 없다. 호기성 조건에서 대장균은 두 개의 서로 다른 말단 퀴놀 산화효소(모두 양성자 펌프)를 사용하여 산소(O2)를 물(H2O)로 환원시킨다.
세균의 복합체 IV는 최종 전자수용체로 역할하는 분자에 따라 분류될 수 있다. 클래스 I 산화효소는 사이토크롬 산화효소이며 산소를 최종 전자수용체로 사용한다. 클래스 II 산화효소는 퀴놀 산화효소이며 다양한 최종 전자 수용체를 사용할 수 있다. 이 두 클래스는 포함하고 있는 산화환원 활성 성분에 따라 세부적으로 분류될 수 있다. 예를 들어 헴 aa3 클래스 I 말단 산화효소는 클래스 II 말단 산화효소보다 훨씬 더 효율적이다.
산소를 최종 전자수용체로 사용하지 않는 혐기성 세균은 최종 전자수용체에 개별화된 말단 환원효소를 가지고 있다. 예를 들어 대장균은 환경에서 이러한 전자 수용체의 가용성에 따라 푸마르산 환원효소, 질산 환원효소, 아질산 환원효소, DMSO 환원효소, 트라이메틸아민-N-옥사이드 환원효소를 사용할 수 있다.
대부분의 말단 산화효소와 환원효소는 유도가능하다. 이들은 특정 환경 조건에 대응하여 필요에 따라 생물에 의해 합성된다.
전자수용체
다양한 전자공여체(유기영양생물의 유기물, 무기영양생물의 무기물)가 있는 것처럼 유기물과 무기물 모두에서 다양한 전자수용체가 있다. 산소를 사용할 수 있는 경우 호기성 세균 및 통성혐기성 미생물은 깁스 자유 에너지 변화량이 가장 크고 가장 많은 에너지를 생성할 수 있기 때문에 산소(O2)를 최종 전자수용체로 사용한다.
대부분의 혐기성 환경에서는 질산염, 아질산염, 제2철, 황산염, 이산화 탄소 및 푸마르산과 같은 유기 분자들을 포함한 다양한 전자수용체가 사용된다.
광합성

산화적 인산화에서 전자는 전자전달계를 통해 NADH와 같은 고에너지 전자공여체로부터 산소(O2)와 같은 전자수용체로 전달된다. 광인산화에서 태양의 빛에너지는 고에너지 전자공여체를 생성하는 데 사용되며, 이는 나중에 산화환원반응의 활성 성분을 환원시킬 수 있다. 전자전달 과정과 화학삼투에 의한 ATP의 합성은 막을 경계로 한 양성자의 농도 기울기에 의해 짝지어진다.
미토콘드리아의 전자전달계와 마찬가지로 광합성의 전자전달계는 세균으로부터 유래된 특별한 시스템으로 간주될 수 있다. 광합성의 전자전달계는 이동성 지용성 퀴논 운반체(필로퀴논 및 플라스토퀴논)와 이동성 수용성 운반체(사이토크롬)을 사용한다. 이들은 또한 양성자 펌프를 포함하고 있다. 모든 광합성이 전자전달계의 양성자 펌프는 미토콘드리아의 복합체 III와 유사하다. 일반적으로 알려져 있는 세포내 공생설은 엽록체와 미토콘드리아가 세균으로부터 유래한 것이라고 설명한다.
같이 보기
더 읽을거리
- Fenchel T, King GM, Blackburn TH (September 2006). 《Bacterial Biogeochemistry: The Ecophysiology of Mineral Cycling》 2판. Elsevier. ISBN 978-0-12-103455-9.
- Lengeler JW (January 1999). Drews G; Schlegel HG, 편집. 《Biology of the Prokaryotes》. Blackwell Science. ISBN 978-0-632-05357-5.
- Nelson DL, Cox MM (April 2005). 《Lehninger Principles of Biochemistry》 4판. W. H. Freeman. ISBN 978-0-7167-4339-2.
- Nicholls DG, Ferguson SJ (July 2002). 《Bioenergetics 3》. Academic Press. ISBN 978-0-12-518121-1.
- Stumm W; Morgan JJ (1996). 《Aquatic Chemistry》 3판. John Wiley & Sons. ISBN 978-0-471-51185-4.
- Thauer RK, Jungermann K, Decker K (March 1977). “Energy conservation in chemotrophic anaerobic bacteria”. 《Bacteriological Reviews》 41 (1): 100–80. doi:10.1128/MMBR.41.1.100-180.1977. PMC 413997. PMID 860983.
- White D (September 1999). 《The Physiology and Biochemistry of Prokaryotes》 2판. Oxford University Press. ISBN 978-0-19-512579-5.
- Voet D, Voet JG (March 2004). 《Biochemistry》. 《Biochemical Education》 28 3판 (John Wiley & Sons). 124쪽. doi:10.1016/s0307-4412(00)00032-7. ISBN 978-0-471-58651-7. PMID 10878303.
- Kim HS, Patel K, Muldoon-Jacobs K, Bisht KS, Aykin-Burns N, Pennington JD, 외. (January 2010). “SIRT3 is a mitochondria-localized tumor suppressor required for maintenance of mitochondrial integrity and metabolism during stress”. 《Cancer Cell》 17 (1): 41–52. doi:10.1016/j.ccr.2009.11.023. PMC 3711519. PMID 20129246.
- Raimondi V, Ciccarese F, Ciminale V (January 2020). “Oncogenic pathways and the electron transport chain: a dangeROS liaison”. 《Br J Cancer》 122 (2): 168–181. doi:10.1038/s41416-019-0651-y. PMC 7052168. PMID 31819197.