

시트르산 회로(영어: citric acid cycle) 또는 TCA 회로(영어: tricarboxylic acid cycle) 또는 크렙스 회로(영어: Krebs cycle)는 세포 호흡의 중간 과정 중 하나로 산소 호흡을 하는 생물에서 탄수화물, 지방, 단백질, 같은 호흡 기질을 분해해서 얻은 아세틸-CoA를 CO2로 산화시키는 과정에서 방출되는 에너지를 ATP(또는 GTP)에 일부 저장하고, 나머지 에너지를 NADH + H+, FADH2에 저장하는 일련의 화학 반응이다. 생성된 NADH + H+, FADH2는 전자전달계로 전달되어 산화적 인산화로 ATP를 생성하는데 사용된다. 시트르산 회로에서 최초로 생성되는 화합물을 정확하게 모르던 시절에 이 화합물이 3개의 카복실기를 가지고 있어서 트라이카복실산 회로(tricarboxylic acid cycle, TCA cycle)라고 이름이 붙여졌었다. 후에 이 트라이카복실산이 시트르산(구연산)이라는 것이 밝혀져서 시트르산 회로로 명명되었고, 발견자인 한스 크렙스의 이름을 따서 크렙스 회로라고도 불린다. 현재 대한민국의 고등학교 생명과학 II 교과서에서는 TCA 회로라고 표기되고 있지만, 대학교의 생명과학 개론서, 생화학 교재, 해외 원서에서는 시트르산 회로로 표기하는 경우가 더 많아서 이 문서에서도 시트르산 회로로 표기하도록 한다. 시트르산 회로는 다른 생화학 반응에서 사용되는 환원제인 NADH, 아미노산 전구체의 공급원이기도 하다. 많은 생화학적 경로에 대해서 시트르산 회로가 가지는 중요성은 시트르산 회로가 생물에서 가장 먼저 확립된 물질대사 체계 중 하나임을 시사한다.
시트르산 회로의 최초 반응은 아세틸-CoA와 옥살아세트산의 축합 반응에 의해 시트르산이 생성되는 것으로 최초 생성물의 이름을 따서 시트르산 회로로 명명되었다. 여기서 시트르산은 citric acid의 이온화 형태인 citrate로 생물체 내의 pH에서는 이온화된 형태가 우세하다. 여기서 논의되는 다른 ~산들도 생물체 내에서 ~ic acid(-COOH)가 아닌 ~ate(-COO−)의 형태로 주로 존재한다. 시트르산 회로는 아세트산(아세틸-CoA의 아세틸기 부분) 및 물을 소비해서 NAD+를 NADH로 환원시키고 부산물로 이산화 탄소를 생성한다. 시트르산 회로에 의해 생성된 NADH는 산화적 인산화 경로로 공급된다. 시트르산 회로와 산화적 인산화를 통해 영양소를 산화시켜 ATP 형태의 사용가능한 화학에너지를 생성한다.
진핵생물에서 시트르산 회로는 미토콘드리아 기질에서 일어나고, 원핵생물에서 시트르산 회로는 세포질에서 일어난다.
발견
시트르산 회로의 여러 구성 요소와 반응들은 센트죄르지 얼베르트의 연구에 의해 1930년대에 밝혀졌다. 센트죄르지 얼베르트는 시트르산 회로의 주요 구성 요소인 푸마르산에 관한 발견으로 1937년에 노벨 생리학·의학상을 수상했다. 센트죄르지는 비둘기 가슴 근육을 이용해서 이러한 발견을 하였다. 비둘기의 가슴 근육 조직은 분쇄기에서 분해되어 수용액에 두어도 산화능력을 잘 유지하기 때문에 산화반응 연구에 매우 적합하다. 시트르산 회로 전체는 셰필드 대학교의 한스 아돌프 크렙스와 윌리엄 아서 존슨에 의해 1937년에 마침내 확인되었고, 이 공로로 1953년에 노벨 생리학·의학상을 수상했다. 시트르산 회로는 발견자인 한스 아돌프 크렙스의 이름을 따서 "크렙스 회로"라고도 불린다.
진화
시트르산 회로의 구성 요소는 혐기성 세균에서 유래되었으며, 시트르산 회로 자체가 한 번 이상 진화했을 수도 있다. 이론적으로 시트르산 회로에 대한 몇 가지 대안이 존재하지만, 시트르산 회로가 가장 효율적인 것으로 보인다. 만약 시트르산 회로에 대한 여러 대안들이 독자적으로 진화해왔다면, 이들은 모두 시트르산 회로에 수렴한 것으로 보여진다.
개요
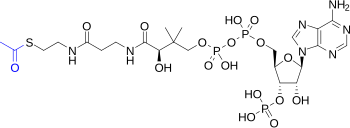
시트르산 회로는 탄수화물, 지방, 단백질 대사를 연결하는 주요 대사 경로이다. 시트르산 회로의 반응들은 아세틸-CoA 형태의 아세트산(C2)을 2분자의 이산화 탄소로 완전히 산화시키는 8가지 효소에 의해 수행된다. 탄수화물, 지방, 단백질의 이화 작용을 통해 생성된 아세틸-CoA가 시트르산 회로로 들어간다. 또한 시트르산 회로 1회전당 3분자의 NADH, 1분자의 FADH2, 기질수준 인산화로 1분자의 GTP(또는 ATP)가 생성된다. 시트르산 회로에 의해 생성된 NADH와 FADH2는 산화적 인산화에 의해 ATP를 생성하는데 사용된다.
해당과정을 통한 탄수화물의 분해로 생성된 피루브산은 아세틸-CoA의 주요 공급원들 중 하나이다. 다음 반응식에 따라 피루브산은 피루브산 탈수소효소 복합체에 의해 산화적 탈카복실화되어 아세틸-CoA를 생성한다.
- 피루브산 + CoA + NAD+ → 아세틸-CoA + NADH + CO2
이 반응의 생성물인 아세틸-CoA는 시트르산 회로의 시작점이다. 또한 아세틸-CoA는 지방산의 산화로부터 얻을 수도 있다. 다음은 시트르산 회로에 대한 개략적인 설명이다.
- 시트르산 회로는 아세틸-CoA로부터 2탄소 아세틸기를 4탄소 수용체 화합물(옥살아세트산)과 축합하여 6탄소 화합물(시트르산)을 형성하는 것으로 시작한다.
- 그런 다음 시트르산은 두 개의 카복시기를 CO2의 형태로 잃어버리는 일련의 화학적 변형을 거친다. 첫 번째 회전에서 CO2로 소실되는 탄소의 기원은 아세틸-CoA가 아닌 옥살아세트산의 탄소이다. 아세틸-CoA의 탄소는 첫 번째 회전에서 CO2로 방출되지 않는다. 아세틸-CoA에 의해 공급된 탄소는 시트르산 회로의 첫 번째 회전 후에 옥살아세트산의 탄소 골격의 일부가 된다. 아세틸-CoA가 공급한 탄소의 CO2의 형태로의 소실은 시트르산 회로의 여러 번의 회전을 필요로 한다. 그러나 시트르산 회로의 중간생성물들은 다른 분자의 생합성을 위한 전구물질로도 사용되기 때문에 아세틸-CoA가 공급한 탄소가 소실되지 않을 수도 있다.
- 시트르산 회로의 산화 반응 단계에 의해 이용가능한 대부분의 에너지는 고에너지 전자의 형태로 NAD+로 전달되어 NADH를 생성한다. 시트르산 회로로 들어가는 아세틸기(아세틸-CoA의 형태로) 1분자당 3분자의 NADH가 생성된다.
- 또한 석신산의 산화에 의해 전자는 석신산 탈수소효소의 보조 인자인 FAD로 전달되어 FADH2를 생성한다. FADH2의 고에너지 전자는 미토콘드리아 내막의 유비퀴논(Q)에 전달되어 유비퀴놀(QH2)로 환원된다. 유비퀴놀은 전자전달계의 복합체 III의 기질이다.
- 시트르산 회로에서 생성된 NADH 와 FADH2는 산화적 인산화를 통해 1분자의 NADH에 의해 2.5분자의 ATP가, 1분자의 FADH2에 의해 1.5분자의 ATP가 생성된다.
- 각 회전이 끝나면 4탄소의 옥살아세트산이 재생성되고, 시트르산 회로가 계속 돌아간다.
단계
2개의 탄소 원자가 CO2 로 산화되고, 이 과정에서 방출된 에너지 중 일부는 GTP(또는 ATP)에 저장되고, 다른 일부는 NADH와 FADH2의 고에너지 전자로 저장된다. 시트르산 회로에서 생성된 NADH는 산화적 인산화라 불리는 과정에서 ATP 생성을 위해 전자를 제공할 수 있다. FADH2는 시트르산 회로와 산화적 인산화가 일어나는 미토콘드리아 내막의 전자전달계에 둘 다 작용하는 효소인 석신산 탈수소효소에 공유 결합되어 있다. 따라서 FADH2는 전자전달계에서 중간생성물로 작용하고, 석신산:유비퀴논 산화환원효소 복합체에 의해 촉매되는 반응의 최종 전자수용체인 유비퀴논(조효소 Q)로의 전자전달을 촉진한다.
시트르산 회로는 아세틸-CoA의 형태로 새로운 탄소를 계속해서 공급받고, 아래 표의 단계 0에서 탄소가 들어간다.
| 단계 (순환) | 기질 | 생성물 | 효소 | 반응 종류 | 설명 |
|---|---|---|---|---|---|
| 0 (10) |
옥살아세트산 + 아세틸-CoA + H2O |
시트르산 + CoA |
시트르산 생성효소 | 알돌 축합 | 비가역적인 반응, 4탄소의 옥살아세트산과 2탄소의 아세틸기가 축합해서 6탄소의 시트르산이 생성됨. |
| 1 | 시트르산 |
시스-아코니트산 + H2O |
아코니테이스 | 탈수 | 가역적인 이성질화 반응 |
| 2 |
시스-아코니트산 + H2O |
아이소시트르산 | 탈수 | ||
| 3 | 아이소시트르산 + NAD+ |
옥살로석신산 + NADH + H+ |
아이소시트르산 탈수소효소 | 산화 | NADH를 생성함 |
| 4 | 옥살로석신산 |
α-케토글루타르산 + CO2 |
탈카복실화 | 속도 제한 단계, 비가역적인 반응, 5탄소의 α-케토글루타르산의 생성 |
|
| 5 |
α-케토글루타르산 + NAD+ + CoA |
석시닐-CoA + NADH + H+ + CO2 |
α-케토글루타르산 탈수소효소 복합체 | 산화적 탈카복실화 | 비가역적인 반응, NADH를 생성함, 4탄소 화합물을 생성 (CoA를 제외) |
| 6 | 석시닐-CoA + GDP + Pi |
석신산 + CoA + GTP |
석시닐-CoA 합성효소 | 기질수준 인산화 | GDP→GTP 또는 ADP→ATP, 1분자의 GTP(또는 ATP)를 생성함 |
| 7 |
석신산 + 유비퀴논 (Q) |
푸마르산 + 유비퀴놀 (QH2) |
석신산 탈수소효소 | 산화 | 석신산 탈수소효소는 보결분자단으로 FAD를 사용하고 반응의 첫 단계에서 FADH2를 생성한다. |
| 8 |
푸마르산 + H2O |
말산 | 푸마레이스 | 수화 | C-C 이중 결합의 수화 |
| 9 |
말산 + NAD+ |
옥살아세트산 + NADH + H+ |
말산 탈수소효소 | 산화 | 가역적인 반응(실제로 평형은 말산쪽으로 기울어져 있다), NADH를 생성함 |
| 10 (0) |
옥살아세트산 + 아세틸-CoA + H2O |
시트르산 + CoA |
시트르산 생성효소 | 알돌 축합 | 이 단계는 단계 0과 동일하며 회로를 다시 시작한다.
비가역적인 반응으로 4탄소의 옥살아세트산과 2탄소의 아세틸기가 축합해서 6탄소의 시트르산이 생성됨. |
사람을 포함한 동물의 미토콘드리아는 두 종류의 석시닐-CoA 합성효소를 갖고 있다. 하나는 GDP로부터 GTP를 생성하고, 다른 하나는 ADP로부터 ATP를 생성한다. 식물은 ATP를 생성하는 종류(ADP-형성 석시닐-CoA 합성효소)를 가지고 있다. 시트르산 회로의 몇몇 효소들은 미토콘드리아 기질 내에서 여러 개의 효소로 구성된 단백질 복합체로 느슨하게 결합될 수 있다.
GDP-형성 석시닐-CoA 합성효소에 의해 생성된 GTP는 뉴클레오타이드 이인산 키네이스(촉매 반응은 GTP + ADP → GDP + ATP 이다)에 의해 ATP를 생성하는데 이용될 수 있다.
생성물
시트르산 회로가 1번 돌 때 1분자의 GTP(또는 ATP), 3분자의 NADH, 1분자의 FADH2, 2분자의 CO2가 생성된다.
1분자의 포도당은 2분자의 아세틸-CoA를 생성하므로 포도당 1분자당 2번의 시트르산 회로가 진행된다. 2번의 시트르산 회로에서 총 2GTP(또는 2ATP), 6NADH, 2FADH2, 4CO2가 생성된다.
| 설명 | 반응물 | 생성물 |
|---|---|---|
| 시트르산 회로의 전체 반응식 | 아세틸-CoA + 3 NAD+ + FAD + Q + GDP + Pi + 2 H2O | → CoA-SH + 3 NADH + FADH2 + 3 H+ + QH2 + GTP + 2 CO2 |
| 피루브산의 산화와 시트르산 회로를 합친 전체 반응식 | 피루브산 + 4 NAD+ + FAD + Q + GDP + Pi + 2 H2O | → 4 NADH + FADH2 + 4 H+ + QH2 + GTP + 3 CO2 |
| 위의 반응식에 해당과정을 합친 전체 반응식 | 포도당 + 10 NAD+ + FAD + 2 Q + 2 ADP + 2 GDP + 4 Pi + 2 H2O | → 10 NADH + 2FADH2 + 10 H+ + 2 QH2 + 2 ATP + 2 GTP + 6 CO2 |
위의 반응식에서 Pi는 H2PO4−이온을 나타내고, ADP는 ADP2−, GDP는 GDP2−, ATP는 ATP3−, GTP는 GTP3−를 각각 나타낸다.
포도당 1분자가 해당과정, 시트르산 회로, 산화적 인산화를 통해 완전히 산화된 후에 얻을 수 있는 ATP의 총 수는 30~38 사이인 것으로 추산된다.
효율성
해당과정, 시트르산 회로, 산화적 인산화에서 포도당 1분자의 산화를 통해 얻을 수 있는 ATP의 이론적인 최대 수율은 38ATP이다(단, 1분자의 NADH당 3ATP, 1분자의 FADH2당 2ATP를 합성한다고 가정했을 경우). 진핵생물에서는 세포질에서 일어나는 해당과정에서 2NADH가 생성된다. 2NADH의 세포질에서 미토콘드리아로의 수송은 2ATP를 소비하므로, ATP의 순생산량을 36으로 감소시킨다. 게다가 미토콘드리아 막을 통한 H+의 누출과 ATP 생성효소, H+펌프의 손실량으로 인한 산화적 인산화에서의 비효율성은 일반적으로 NADH와 FADH2로부터의 ATP 수율을 이론적인 최대 수율보다 낮춘다. 따라서 관찰된 수율은 NADH당 약 2.5ATP와 FADH2당 약 1.5ATP에 더 가깝고, ATP의 총 순생산량을 약 30ATP로 더 감소시킨다. 새로 수정된 H+ 대 ATP의 비율로 총 ATP 수율을 계산하면 포도당 1분자당 29.85ATP의 추정치가 나온다.
변이
시트르산 회로는 일반적으로 잘 보존되어 있지만, 다른 분류군에서 발견되는 효소들에서 상당한 변이가 있다 (이 문서의 도표는 포유류의 대사 경로 변이를 다룬다).
진핵생물과 원핵생물 간에는 약간의 차이가 존재한다. 아이소시트르산을 α-케토글루타르산으로 전환시키는 반응을 촉매하는 아이소시트르산 탈수소효소는 진핵생물에서는 NAD+ 의존적인 EC 1.1.1.41 이고, 원핵생물에서는 NADP+ 의존적인 EC 1.1.1.42이다. 비슷하게 말산을 옥살아세트산으로 전환시키는 반응을 촉매하는 말산 탈수소효소는 진핵생물에서는 NAD+ 의존적인 EC 1.1.1.37 이고, 대부분의 원핵생물에서는 퀴논 의존적인 EC 1.1.5.4이다.
석시닐-CoA를 석신산으로 전환하는 석시닐-CoA 합성효소가 촉매하는 단계로 상당한 변이가 있는 단계이다. 대부분의 생물들은 EC 6.2.1.5, 석신산-CoA 연결효소 (ADP-형성) (그 이름에도 불구하고 효소는 ATP 생성 방향으로 작용한다)를 사용한다. 포유류에서는 GTP를 생성하는 효소인 석신산-CoA 연결효소 (GDP-형성) (EC 6.2.1.4) 도 사용한다. 각 동질효소의 사용 정도는 조직에 따라 다르다.Acetobacter aceti 와 같은 일부 아세트산 생성 세균에서는 완전히 다른 효소(EC 2.8.3.18, 석시닐-CoA : 아세트산 CoA 전이효소)가 이 반응을 촉매한다. 이러한 특이 효소들은 이들 아세트산 생성 세균에서 시트르산 회로와 아세트산 대사를 연결시킨다.Helicobacter pylori 와 같은 일부 세균은 이 과정에 석시닐-CoA : 아세토아세트산 CoA 전이효소(EC 2.8.3.5)라는 또 다른 효소를 사용한다.
또한 α-케토글루타르산의 석시닐-CoA로의 전환 단계에도 몇몇 동질효소가 존재한다. 대부분의 생물은 NAD+ 의존성 α-케토글루타르산 탈수소효소를 사용하지만, 일부 세균은 페레독신 의존성 α-케토글루타르산 생성효소(EC 1.2.7.3)을 사용한다. 독립 영양 세균, 메테인 생성균, 고세균을 포함하는 다른 생물들은 석시닐-CoA를 완전히 우회해서 α-케토글루타르산을 석신산 세미알데하이드를 거쳐 석신산으로 전환시키는 반응을 α-케토글루타르산 탈카복실화효소(EC 4.1.1.71), 석신산-세미알데하이드 탈수소효소(EC 1.2.1.79)를 통해 수행한다.
조절
시트르산 회로의 조절은 주로 생성물에 의한 억제와 기질의 이용가능성에 의해 결정된다. 만약 시트르산 회로가 조절되지 않은 채로 돌아가면, NADH와 같은 환원된 조효소와 ATP의 과잉 생산으로 많은 양의 대사 에너지가 낭비될 수 있다. 시트르산 회로의 중요한 조절 물질은 ATP로 전환되는 ADP이다. ADP 양의 감소는 회로의 여러 효소들을 차례로 억제할 수 있는 전구물질인 NADH의 축적을 야기한다. 석신산 탈수소효소를 제외한 시트르산 회로의 모든 탈수소효소의 생성물인 NADH는 피루브산 탈수소효소, 아이소시트르산 탈수소효소, α-케토글루타르산 탈수소효소, 시트르산 생성효소를 억제한다. 아세틸-CoA는 피루브산 탈수소효소를 억제하고, 석시닐-CoA는 α-케토글루타르산 탈수소효소와 시트르산 생성효소를 억제한다. 시험관에서 시트르산 회로 효소로 실험할 때 ATP는 시트르산 생성효소와 α-케토글루타르산 탈수소효소를 억제한다. 그러나 생체 내에서 휴식과 격렬한 운동 사이에 ATP의 농도는 10% 이상 변하지 않는다. 농도 변화가 10% 미만으로 변하는 다른 자리 입체성(알로스테릭) 효과 인자로 반응 속도의 큰 변화를 설명할 수 있는 다른 자리 입체성 조절 메커니즘은 알려져 있지 않다.
또한 칼슘도 시트르산 회로에서 조절 인자로 사용된다. 미토콘드리아 기질에서 칼슘 수치는 세포가 활성화될 때 수 십 마이크로몰 수준까지 이를 수 있다. 이는 피루브산 탈수소효소 포스파테이스를 활성화시켜 피루브산 탈수소효소 복합체를 활성화시킨다. 또한 칼슘은 아이소시트르산 탈수소효소와 α-케토글루타르산 탈수소효소를 활성화시킨다. 이것은 시트르산 회로에서 많은 단계의 반응 속도를 증가시키므로, 경로 전체의 흐름이 증가한다.
시트르산은 피루브산의 전구물질인 과당 1,6-이중인산의 형성을 촉매하는 해당과정의 효소인 포스포프럭토키네이스를 억제하기 때문에 피드백 억제에 이용된다. 이는 시트르산이 축적되고 효소에 대한 기질이 감소할 때 대사 흐름의 속도가 높게 유지되는 것을 방지한다.
최근의 연구는 시트르산 회로의 중간생성물과 저산소증 유도인자(HIF, hypoxia-inducible factor)의 조절 간에 중요한 연관성이 있음을 보여주었다. 저산소증 유도인자는 산소 공급이 제한적일 때 혈관 형성, 혈관 재구성, 포도당 이용, 철분 수송 및 세포 자살에 작용하는 전사인자이다. 저산소증 유도인자에서 두 개의 중요한 프롤린 잔기 중 적어도 하나의 하이드록실화는 E3 유비퀴틴 연결효소와의 상호작용을 매개한다. 이 반응은 프롤린 하이드록실레이스에 의해 촉매된다. 푸마르산 및 석신산은 프롤린 하이드록실레이스의 강력한 억제제로 저산소증 유도인자의 안정화를 유도한다.
시트르산 회로로 수렴하는 주요 대사경로
몇몇 이화작용 경로가 시트르산 회로로 수렴한다. 이러한 반응의 대부분은 중간생성물을 시트르산 회로로 첨가하기 때문에 보충대사 반응(anaplerotic reaction)으로 알려져 있다. 이것은 시트르산 회로로 들어올 수 있는 아세틸-CoA의 양을 증가시켜서 호흡을 수행하는 미토콘드리아의 능력을 증가시킨다. 시트르산 회로에서 중간생성물을 제거하는 과정은 "cataplerotic" 반응이라고 한다.
해당과정에서 생성된 피루브산은 미토콘드리아 내막을 가로질러 미토콘드리아 기질로 능동수송된다. 미토콘드리아 기질에서 피루브산은 산화적 탈카복실화되고, 조효소 A(CoA)와 결합하여 아세틸-CoA, NADH, CO2를 형성한다.
그러나 피루브산은 피루브산 카복실화효소에 의해 카복실화되어 옥살아세트산을 형성하는 것도 역시 가능하다. 피루브산 카복실화효소에 의한 반응은 보충대사 반응으로 시트르산 회로의 옥살아세트산의 양을 채우고, 활동으로 인해 조직(예: 근육에서)의 에너지 요구량이 증가할 때 시트르산 회로의 아세틸-CoA 대사 능력을 증가시킨다.
시트르산 회로에서 모든 중간생성물들(예: 시트르산, 아이소시트르산, α-케토글루타르산, 석시닐-CoA, 석신산, 푸마르산, 말산, 옥살아세트산)은 회로가 도는 동안 재생성된다. 따라서 시트르산 회로의 중간생성물들 중에 하나를 더 많이 미토콘드리아에 첨가해주면 회로 내 다른 중간생성물로 차례로 전환되어 다른 모든 중간생성물의 양이 증가하므로 추가된 양이 시트르산 회로 내에서 유지된다. 따라서 시트르산 회로에 중간생성물들 중 하나를 추가하면 "anaplerotic" 효과가 나타나고, 중간생성물들 중 하나를 제거하면 "cataplerotic" 효과가 나타난다. 이러한 보충대사(anaplerotic) 반응 및 cataplerotic 반응은 시트르산 회로 주기 동안 아세틸-CoA와 결합하여 시트르산을 형성할 수 있는 옥살아세트산의 양을 증가시키거나 감소시킨다. 이것은 차례로 미토콘드리아에 의한 ATP의 생성 속도를 증가시키거나 감소시켜서, 세포의 ATP 이용가능성을 증가 또는 감소시킨다.
한편 피루브산의 산화 또는 지방산의 β 산화로부터 생성된 아세틸-CoA는 시트르산 회로로 들어간다. 시트르산 회로의 각 회전마다 아세틸-CoA는 옥살아세트산과 반응하여 시트르산으로 축합되고, 다시 아세틸-CoA로 재생성되진 않는다. 아세틸-CoA의 아세틸기 부분이 산화되어 CO2와 H2O로 분해되고, 이 과정에서 방출되는 에너지의 일부는 열에너지로 소실되고 일부는 ATP에 화학에너지의 형태로 저장된다. 지방산의 β 산화에서 세 단계 과정은 시트르산 회로에서 석신산이 옥살아세트산으로의 전환에서 일어나는 반응들과 유사하다. 지방산의 β 산화에서 FAD가 FADH2로 환원되는 동안 아세틸-CoA는 트랜스-엔오일-CoA로 산화된다. 이는 시트르산 회로에서 석신산이 푸마르산으로 산화되는 반응과 유사하다. 다음으로 β 산화에서 트랜스-엔오일-CoA는 이중결합을 가로질러 수화되어 β-하이드록시아실-CoA가 되는 반응은 시트르산 회로에서 푸마르산이 말산으로 수화되는 반응과 유사하다. 마지막으로 β-하이드록시아실-CoA는 β-케토아실-CoA로 산화되고, NAD+는 NADH로 환원되는 반응은 시트르산 회로에서 말산을 옥살아세트산으로 산화시키는 것과 동일한 과정을 따른다.
혈액 내 글루카곤 또는 에피네프린의 농도가 증가시 H 간에서 세포질의 피루브산의 미토콘드리아 내의 옥살아세트산으로의 카복실화는 젖산 및 탈아미노화된 알라닌을 포도당으로 전환시키는 포도당신생합성의 초기 반응 단계이다. 여기서 미토콘드리아에 옥살아세트산을 첨가하면 시트르산 회로의 또 다른 중간생성물인 말산이 즉각적으로 미토콘드리아에서 제거되어 세포질의 옥살아세트산으로 전환되기 때문에 보충대사(anaplerotic) 효과가 일어나지 않는다. 이러한 포도당신생합성은 해당과정의 거의 역반응으로 이들은 결국 포도당으로 전환된다.
단백질의 분해에서 단백질은 단백질 가수분해효소(protease)에 의해 아미노산으로 분해된다. 이들의 탄소 골격(즉, 탈아미노화된 아미노산)은 시트르산 회로에 대해 보충대사(anaplerotic) 효과를 갖는 중간생성물들(예: 글루탐산 또는 글루타민으로부터 유도된 α-케토글루타르산)로 시트르산 회로로 유입될 수 있다. 또는 류신, 아이소류신, 라이신, 페닐알라닌, 트립토판, 티로신, 트레오닌의 경우 아세틸-CoA로 전환되어 CO2와 H2O로 산화되거나 케톤체를 형성하는데 사용된다. 케톤체는 간이 아닌 다른 조직에서만 산화되거나 또는 소변이나 호흡을 통해 배설될 수 있다. 중간생성물로서 시트르산 회로로 유입되는 아미노산들은 미토콘드리아 밖으로 수송되는 말산이 세포질의 옥살아세트산으로 전환되어 궁극적으로 포도당이 만들어지는 포도당생성(gluconeogenic) 아미노산들로 시트르산 회로에서 유출될 수 있다. 반면에 아세토아세틸-CoA나 아세틸-CoA를 거쳐 케톤체를 생성하는 아미노산들(류신, 아이소류신, 라이신, 페닐알라닌, 트립토판, 타이로신, 트레오닌)은 케톤생성(ketogenic) 아미노산이다. 탈아미노화된 알라닌, 시스테인, 글리신, 세린, 트레오닌, 트립토판은 피루브산으로 전환되고 옥살아세트산으로(보충대사 반응으로) 시트르산 회로로 유입되거나 또는 피루브산에서 아세틸-CoA로 전환된 다음 CO2와 H2O로 분해될 수 있다.
지방의 분해에서 트라이글리세라이드(중성 지방)가 가수분해되어 지방산과 글리세롤로 전환된다. 간에서 글리세롤은 포도당신생합성에서 다이하이드록시아세톤 인산과 글리세르알데하이드 3-인산을 거쳐 포도당으로 전환될 수 있다. 많은 조직들에서 특히 심장 및 골격근에서 지방산은 β 산화로 알려진 과정을 통해 분해된다. β 산화는 시트르산 회로에서 사용될 수 있는 아세틸-CoA를 생성한다. 탄소 수가 홀수 개인 지방산의 β 산화는 프로피오닐-CoA를 생성하고, 다음에 석시닐-CoA로 전환되어 보충대사(anaplerotic) 반응의 중간생성물로 시트르산 회로로 유입된다.
1분자의 NADH 산화에 의해 약 2.5분자의 ATP, 1분자의 FADH2의 산화에 의해 약 1.5분자의 ATP가 합성된다고 하면, 1분자의 포도당이 해당과정(2ATP, 2NADH), 피루브산의 산화(2NADH), 시트르산 회로(2ATP, 6NADH, 2FADH2), 산화적 인산화(28ATP)에 의해 최대 32분자의 ATP가 생성될 수 있다.
시트르산 회로의 중간생성물은 생합성 과정에 기질로 사용됨
시트르산 회로의 몇몇 중간생성물들은 주요 화합물의 합성에 사용된다. 아세틸-CoA는 미토콘드리아 밖으로 수송될 수 없다. 세포질의 아세틸-CoA를 얻기 위해, 시트르산 회로로부터 시트르산이 유출되고, 시트르산은 미토콘드리아 내막을 가로질러 세포질로 운반된다. 시트르산은 세포질에서 ATP 시트르산 분해효소(ATP citrate lyase)에 의해 아세틸-CoA와 옥살아세트산으로 분해된다. 옥살아세트산은 말산으로 전환된 다음 미토콘드리아 기질로 다시 들어가고 더 많은 아세틸-CoA를 미토콘드리아 밖으로 운반하기 위해 옥살아세트산으로 다시 전환된다. 세포질의 아세틸-CoA는 지방산의 합성 및 콜레스테롤 합성에 사용된다. 콜레스테롤은 스테로이드 호르몬, 담즙산, 비타민 D를 합성하는데 사용될 수 있다.
많은 비필수 아미노산의 탄소 골격은 시트르산 회로의 중간생성물로부터 만들어진다. 시트르산 회로의 중간생성물로부터 형성된 α-케토산을 아미노산으로 전환시키기 위해 피리독살 인산이 보조 인자인 아미노기 전이반응에서 글루탐산의 아미노기를 α-케토산으로 전이시킨다. 이 반응에서 글루탐산은 시트르산 회로의 중간생성물인 α-케토글루타르산으로 전환된다. 아미노산 합성을 위해 탄소 골격을 제공할 수 있는 중간생성물에는 아스파르트산, 아스파라긴을 형성하는 옥살아세트산과 글루타민, 프롤린, 아르지닌을 형성하는 α-케토글루타르산이 있다.
이들 아미노산들 중 아스파르트산과 글루타민은 ATP, AMP, GTP, NAD+, FAD, CoA 뿐만 아니라 DNA와 RNA의 염기로 사용되는 퓨린을 만드는데 사용된다.
피리미딘은 부분적으로 아스파르트산(옥살아세트산으로부터 유도됨)으로부터 만들어진다. 피리미딘인 티민, 사이토신, 유라실은 DNA 및 RNA에서 퓨린 염기의 상보적인 염기로 작용하며, CTP, UMP, UDP, UTP의 구성 요소이다.
포르피린의 탄소 원자의 대부분은 시트르산 회로의 중간생성물인 석시닐-CoA에서 유래한다. 포르피린은 헤모글로빈, 미오글로빈, 다양한 사이토크롬과 같은 헴단백질의 주요 구성 요소이다.
포도당신생합성 동안 미토콘드리아의 옥살아세트산은 말산으로 환원된 다음 미토콘드리아 밖으로 운반되고, 세포질에서 옥살아세트산으로 다시 산화된다. 세포질의 옥살아세트산은 포스포엔올피루브산 카복시키네이스에 의해 포스포엔올피루브산으로 탈카복실화된다. 포스포엔올피루브산 카복시키네이스가 촉매하는 반응은 간과 콩팥에서 거의 모든 포도당신생합성 전구물질들(포도당생성 아미노산과 젖산과 같은)을 포도당으로 전환하는 속도 제한 단계이다.
따라서 시트르산 회로는 이화작용과 동화작용에 모두 관여한다.
순환하는 젖산을 통해 포도당은 시트르산 회로로 공급됨
젖산의 대사 과정에서의 역할은 조직과 종양에서 연료로 사용된다는 것으로 잘 알려져 있다. 코리 회로에서 근육은 젖산을 생성하고 간으로 흡수된 젖산은 포도당신생합성에 사용된다. 새로운 연구에 따르면 젖산은 시트르산 회로에 탄소 공급원으로 사용될 수 있다.
같이 보기
외부 링크
- An animation of the citric acid cycle
- Citric acid cycle variants
- Pathways connected to the citric acid cycle Archived 2008년 10월 26일 - 웨이백 머신 at Kyoto Encyclopedia of Genes and Genomes
- Introduction at Khan Academy Archived 2014년 8월 25일 - 웨이백 머신
- metpath: Interactive representation of the citric acid cycle
| 옥살아세트산 | 말산 | 푸마르산 | 석신산 | 석시닐-CoA | ||||||||||||

|

|

|

|

|
||||||||||||
|
|
|
|
|
|||||||||||||
| 아세틸-CoA | NADH + H+ |
NAD+ | H2O | FADH2 | FAD | CoA + ATP (GTP) |
Pi+ ADP (GDP) |
|||||||||
|
|
+ | H2O |
|
|
NADH + H++ CO2 |
|||||||||||
| CoA | NAD+ | |||||||||||||||

|
H2O |

|
H2O |

|
NAD(P)+ | NAD(P)H + H+ |

|
CO2 |

|
|||||||
|
|
|
|
|
|||||||||||||
| 시트르산 | 시스아코니트산 | 아이소시트르산 | 옥살석신산 | α-케토글루타르산 | ||||||||||||