
|
| ||
|---|---|---|
| 화석 범위: 지바절 - 현재 | ||
 미국 어류 및 야생동물관리국의 현대 늑대 (사진가:Gary Kramer). | ||
|
🔊 회색늑대의 소리 | ||
| 생물 분류ℹ️ | ||
| 역: | 진핵생물 | |
| 계: | 동물계 | |
| 문: | 척삭동물문 | |
| 아문: | 척추동물아문 | |
| 상강: | 사지상강 | |
| 강: | 포유강 | |
| 하강: | 진수하강 | |
| 목: | 식육목 | |
| 아목: | 개아목 | |
| 과: | 개과 | |
| 아과: | 개아과 | |
| 족: | 개족 | |
| 속: | 개속 | |
| 종: | 회색늑대 | |
| 학명 | ||
| Canis lupus | ||
| Linnaeus, 1758 | ||
| 향명 | ||
| 아종 | ||
|
39개 아종. 본문 참조
| ||
| 회색늑대의 분포 | ||
 | ||
| 보전상태 | ||

회색늑대(학명: Canis lupus 카니스 루푸스[*])는 개과의 동물로 북아메리카, 유라시아, 북아프리카 황무지에 서식하는 가축화된 개의 조상이며 이리나 말승냥이로도 불린다. 늑대는 수컷 평균 43~45 kg, 암컷 평균 36~38.5kg이고 어깨높이는 0.6~0.9m로 개과에 속한 종 중 가장 큰 종이다. 늑대는 썰매개 또는 저먼 셰퍼드와 모양과 비율이 비슷하나, 늑대가 큰 머리, 좁은 가슴, 긴 다리, 똑바로 선 꼬리와 큰 발을 가지고 있는 점이 다르다. 겨울 털은 크고 무성하며 털 색은 일반적으로 얼룩덜룩한 회색이지만 거의 순수한 흰색, 붉은색, 갈색, 검은색 등의 다양한 색도 있다.
늑대의 속은 개속에 속하며, 회색늑대는 더 작은 아속인 코요테나 황금자칼과 비교하여 형태학적으로 몸집이 큰 먹이를 사냥하는 데 갖춰주었고 좀 더 집단적인 성격을 갖추며, 고도의 의사소통을 갖추어서전문종으로 분류한다. 늑대는 사회적 동물로서 한 쌍의 성체와 그 자손으로 구성된 핵가족의 부계사회 형태로 이루어져 있다. 늑대는 최상위 포식자로 천적이 호랑이와 표범, 불곰과 회색곰, 검독수리 그리고 인간밖에 없다. 다만 검독수리의 경우 중소형 개체만을 노리며 대형 개체는 사냥하지 않는다. 늑대는 주로 대형 유제류를 먹이로 삼으나 이 먹이들이 부족한 경우에는 작은 동물, 가축, 썩은 고기, 쓰레기를 먹기도 한다.
회색늑대는 세계에서 가장 잘 연구된 동물 중 하나이며, 다른 야생 동물 종들보다 더 많은 연구 서적이 있다. 늑대는 인간과의 오랜 역사를 가지고 있으며, 보통 농업 사회에서 가축을 공격하는 경멸받는 존재로 인식되어서 사냥되기도 하였지만, 반대로 일부 아메리카 토착민들에게는 존중되기도 하였다.개의 선조는 늑대이며, 북아시아에서 가축화가 시작되었다. 인간 사회에서는 늑대가 중세 늑대의 인간 공격으로 발생한 광견병에 기인하여 두려움의 상징으로 퍼져 있었다. 광견병에 걸리지 않은 늑대가 주로 어린이들을 공격한 적도 있으나, 이는 특이한 경우이고 상대적으로 늑대는 인간 사회와 떨어져서 생활하며 사냥꾼과 목동들로 인해 인간이 두려운 존재로 습득되었다. 늑대는 사냥과 덫으로 인해 종 범위가 3번째로 빨리 감소하였으나, 여전히 광범위한 범위의 서식지와 안정적인 개체 수로 인해 세계적으로 종 감소 위협은 아니라고 인식되어 국제 자연 보호 연맹은 "관심 필요"의 낮은 등급을 부여했다.
어원
영어의 "wolf"라는 단어는 고대 영어의 "wulf"로, 이는 고대 게르만어파 "*wulfaz"에서 파생된 것으로 추측하며 또 이는 고대 인도유럽어족의 "*wlqwos"/"*lukwos"라는 단어에서 파생된 것으로 추측된다.고대 영문학에서는 앵글로색슨 왕과 그 전사 이름의 접두사와 접미사에 "wulf"라는 명칭을 붙였다. 그 예로, Wulfhere, Cynewulf, Ceonwulf, Wulfheard, Eanwulf, Wulfmeer, Wulfstan, Aethelwulf 등의 이름이 있었다. 또한, 늑대와 관련된 이름으로 기독교화 이전 게르만족의 전사들 이름에 Wolfhroc (Wolf-Frock), Wolfhetan (Wolf Hide), Isangrim (Grey Mask), Scrutolf (Garb Wolf), Wolfgang (Wolf Gait) and Wolfdregil (Wolf Runner) 등으로 늑대와 관련된 이름을 붙였다.
라틴어의 lupus는 사비니인의 외래어로에드워드 톱셀에 따르면 원래 이 말은 그리스어로 사자발을 뜻하는 λεοποσ (leopos)과 빛을 뜻하는 λουκᾶς (loukas) 둘 모두로 유도할 수 있다라고 말했다.
늑대는 한국어에서 이리라고도 했으며, 문헌에서 이리(狼)와 늑대(勒大)를 구별하기도 했다. 회색늑대는 영어명을 번역한 이름이나, 한반도에 살던 아종 늑대(C. l. chanco)를 포함하기 위한 이름으로 사용한다.
| 언어 또는 지역어 이름 | 현지어 이름 |
|---|---|
| 아이누어 | Seta |
| 알바니아어 | Ujku |
| 아랍어 | ذئب (the’b) |
| 아라파호어 | Haqihana |
| 카도어 | Tasha |
| 중국어 | 狼 (láng) |
| 콘월어 | Bleit |
| 크리어 | May-hee'-gan |
| 크로아티아어 | Vuk |
| 체코어 | Vlk |
| 덴마크어/노르웨이어 | Ulv |
| 네덜란드어/영어/독일어 | Wolf |
| 에스토니아어 | Hunt, susi |
| 핀란드어 | Susi |
| 프랑스어 | Loup |
| 그리스어 | Λύκος (lukos) |
| 구자라트어 | ના○ (Bagad) |
| 힌디어 | Bheriya, gúng, hondár, nekra, bighána |
| 헝가리어 | Farkas |
| 이누크티투트어 | ᐊᒪᕈᖅ (amaruq) |
| 아일랜드어 | Faol, faol-chu |
| 이탈리아어 | Lupo |
| 일본어 | オオカミ(狼) (ōkami), ヤマイヌ(豺、犲、山犬) (yamainu) |
| 카슈미르어 | Ratnakin |
| 한국어 | 늑대, 이리, 말승냥이 |
| 라다키어 | Chanco |
| 라트비아어 | Vilks |
| 리투아니아어 | Vilkas |
| 몽골어 | ᠴᠢᠨᠤᠠ / Чоно (chono) |
| 나바호어 | Maicoh |
| 누나미우트어 | Amaguk |
| 오글라 라코타어 | Shunk'-ah mah-nee'-tu |
| 오지브웨어 | My-in'-gan, kit'-chi my-in'gan |
| 페르시아어 | گرگ (gorg) |
| 폴란드어 | Wilk |
| 포르투갈어/스페인어 | Lobo |
| 루마니아어 | Lup |
| 러시아어 | Волк (volk) |
| 스코틀랜드 게일어 | Madadh, madadh allaidh |
| 신디어 | Bagyár |
| 슬로바키아어 | Vlk dravý |
| 슬로베니아어 | Volk |
| 스웨덴어 | Varg |
| 텔루구어 | Toralú |
| 튀르키예어 | Kurt |
| 웨일스어 | Bleidd |
| 양콘 수족어 | Soŋ toke cha tuŋ-ka |
진화와 분류


기원

선조
늑대 종의 가장 가능성이 높은 선조 후보는 북아메리카에서 마이오세 시기 서식한 토끼잡이늑대(Canis lepophagus)로 작고 좁은 두개골을 가졌으며 이 모습은 코요테와의 유전적 관계도 설명이 가능했다. 크고 뚱뚱한 보로파구스아과(Borophaginae)가 멸종한 후, 레포파구스는 몸집이 커지고 넓은 두개골을 가진 동물이 되었다. 이러한 형태의 레포파구스는 텍사스 북부에서 발견되었으며 현대의 늑대와 비슷한 모양이었다. 북아메리카에서의 첫 번째 현대 늑대의 발견은 블랜캔(Blancan) 후기와 이르빙토닌(Irvingtonian) 초기이다. 이 토끼잡이늑대 종 중에는 현대의 아메리카붉은늑대와 비슷한 종이 있었으며 아마도 유라시아에서 베링 육교를 건넌 것으로 추측된다. 유라시아의 새로운 프리스콜라트란스(C. priscolatrans) 종은 점차 모스바체시스(C. mosbachensis) 종으로 대체되고아라비아 반도와 서아시아에서는 현대 늑대와 밀접한 닮은 화석이 발견되며, 유럽에서는 늑대가 50만 년 전 빙하기 시기에 퍼졌다(늑대#아종 참조). 모르바체시스는 후에 현대 늑대로 진화한다.
아종화
늑대의 미토콘드리아 DNA 연구에서, 적어도 회색늑대의 뚜렷한 혈통은 가장 오래된 북아프리카, 서아프리카, 동아프리카가 서식지인 아프리카늑대로 플라이스토세 중기에서 말기에 처음 나타난 것으로 추측된다.인도 아대륙에 나타난 또 다른 늑대의 혈통은 카슈미르, 히마찰프라데시 주, 동부 네팔 및 티베트 일부 등 히말라야 동부가 서식지인 티베트늑대로, 이 종은 히말라야 주변의 지질과 기후 대변동이 있는 약 80만 년 전부터 서식하기 시작한 것으로 추측된다. 인도 반도에 서식하는 인도늑대(C. l. pallipes)는 약 40만 년 전 히말라야 늑대에서 갈라져 나왔다. 티벳늑대는 15만 년 전 카슈미르 북서부에서 갈라져 나온 것으로 추측된다. 가장 마지막으로 갈라진 이 혈통은 전북구에서 유럽과 북미로 확장하여 유전자 표지자 그림과 같이 유럽 늑대, 북미 늑대, 개와 비슷한 혈통에 속해 있다.
지금은 멸종한 일본늑대는 일본에 서식한 늑대로 이는 플라이스토세에 2만 년 전 아시아 본토에서 갈라지기 전에 동부 시베리아의 늑대에서 갈라져 나온 것으로 추측된다. 플라이스토세 동안, 쓰가루 해협의 확대로 혼슈와 홋카이도섬이 분리되고 이는 일본 열도에서 서식한 가장 뚱뚱한 우제류의 멸종으로 이어지는 기후 변화가 찾아온다. 일본 늑대는 7천 년에서 13,000년 전 기후학적, 생태학적인 이유로 섬의 왜소 발육화가 찾아온 것으로 추측된다. 홋카이도늑대(C. l. hattai)는 사촌격인 일본늑대(C. l. hodophilax)보다 크기가 크며, 이는 높은 고도에서 살며 큰 먹이를 얻을 수 있는 것뿐 아니라 시베리아 늑대에서 분산된 유전자의 상호 작용으로도 볼 수 있다.
북아메리카 이동
현대 늑대는 란촐라브린(Rancholabrean) 시기 북미로 확장된 것으로 추측된다. 큰 크기의 다이어늑대(Canis dirus)가 이미 북아메리카에 존재했으나, 이미 8천 년 전 멸종했고 큰 사냥감들은 없어졌다. 새로 도착한 회색늑대와의 경쟁에서 작고 빠른 먹이들을 먹이 위해 종 개체 수가 감소하였으나 살아남을 수 있었다. 다이어늑대의 멸종과 함께, 회색늑대는 유일한 대규모 확산적 갯과 종이었다. 북미의 재식민지화의 모습이 여러 곳에서 나오며, 가장 특이한 종들이 주변에서 나오기 시작했다. 이 종들(북극늑대(C. l. arctos)는 북극의 높은 섬에 생존, 동부늑대(C. l. lycaon)는 북아메리카 동부 숲에서, 멕시코늑대(C. l. baileyi)는 멕시코 등지의 남쪽에 서식)은 유라시아에서의 이전 종들의 모습을 볼 수 있었다. 동부늑대와 멕시코늑대는 일부 원시적인 특성과 체계적인 친화성을 가지고 있었다. 캘리포니아 남부 해안의 화석에서 북극늑대와 비슷한 툰드라늑대(C. l. albus)가 플라이스토세 후기에 큰 크기의 모습으로 존재하고 있었고, 북미의 대형 회색늑대의 아종이 남아 있으며 현재 늑대는 더는 그곳에 거주하진 않지만 오래전에는 빙하 주기에 의해 남쪽으로 이주한 것으로 추측된다. 멕시코늑대와 비슷한 작은 늑대가 캔자스와 남부 캘리포니아 등 다양한 지역에서 발견되었다. 이는 플라이스토세 후기 늑대들의 북극 생활에서 남쪽으로 이동하며 유랑 생활을 하며 따뜻한 곳에서 대량 적응한 것으로 보인다.
아종


2005년 기준으로,MSW3에서는 37개의 아종을 인정하고 있다. 이들 가운데는 개와 딩고도 있다.
늑대의 아종은 보통 두 가지 범주로 구성된다:
- "북부늑대": 몸집과 두뇌가 크고, 치아가 열육성이며 북미와 유럽, 북아시아에 서식한다.
- "남부늑대": 아라비아 반도, 남아시아 및 북아프리카에 서식한다. 이들은 두개골과 이빨이 작은 특징이 있으며, 털이 잔털로 짧고 얕다. 이들은 고대 유럽 늑대의 화석과 비슷하게 생겨 초기 늑대의 잔존 생물로 추측된다. 이 늑대들은 DNA 상 미국과 유럽의 15만 년 전 분리된 혈통에 비해 80만 년 전 분리되었다. 남부늑대의 울음소리는 짧고 높은 개 짖는 소리와 비슷하며, 드물게는 울부짖기도 한다(howling). 이들은 개와 딩고의 선조로도 추측된다.
중앙과 동아시아의 늑대들은 북부와 남부늑대의 중간 정도의 특징을 가지고 있다. 이 늑대들의 뇌 크기 차이는 크게 밝혀져 있는데, 가장 큰 크기의 북유라시아의 늑대와 약간 작은 머리를 가진 북미 늑대와 가장 작은 남부늑대(북부늑대보다 5~10% 작음)로 구별할 수 있다.
현재 늑대의 아종은 대략 13~15종류의 아종으로 묶을 수 있다(단, 여기에 제시된 털의 색깔은 절대적이 아니다.)
- 아랍늑대 Canis lupus arabs : 심각한 멸종 위기 상태이며, 지금도 그 숫자가 줄어들고 있다. 남부 이스라엘과 사우디아라비아, 오만, 예멘 등에 걸쳐 서식하고 있다. 아주 작은 체격의 아종이며, 전형적으로 혼합된 갈색 및 완연한 갈색빛을 띠고 있다. 해로운 짐승(해수, 害獸)으로 몰려 사냥당하고 있다.
- 북극늑대 Canis lupus arctos : 비교적 안정적인 상태이며, 캐나다령 북극과 그린란드에 걸쳐 서식한다. 평균 치수가 재진 아종이다. 거의 전체적으로 두꺼운 외투를 가진 흰색 또는 크림색의 흰색이다. 희소하지만 합법적으로 사냥당하고 있다.
- 딩고 Canis lupus dingo : 순수 혈통을 가진 종으로 자칫 멸종위기에 처하기 쉽다. 오스트레일리아와 동남아시아에 서식하고 있다. 합법적으로 사냥당하고 있으며, 순수 혈통은 집 개와의 이종교배(異種交配)로 인해 줄어들고 있다.
- 개 Canis lupus familiaris : 안정적인 상태이며, 전 세계적으로 분포하고 있다.
- 유라시아늑대 Canis lupus lupus : 안정적인 상태이다. 서유럽, 스칸디나비아, 러시아, 몽골 북서부, 중국 북부에서 산다. 평균적으로 큰 체격이며, 일반적으로 별로 길지 않은 혼합된 털빛깔을 띤다.
- 대평원늑대 Canis lupus nubilus : 멸종된 아종이며 미국 대평원에서 1926년에 자취를 감추었다.
- 매켄지계곡늑대 Canis lupus occidentalis : 알래스카 지역, 북부 로키산맥, 서부와 중앙 캐나다에 걸쳐 서식하고 있으며 옐로스톤국립공원에서 서식중인 아종이다.
- 멕시코늑대 Canis lupus baileyi : 심각한 멸종위기 상태의 아종이다. 중부 멕시코, 서부 텍사스, 중부 뉴멕시코와 텍사스에 걸쳐 산다. 비교적 작은 체격의 아종이다. 털빛은 황갈색 계통 및 쇠 녹빛의 갈색이며 등은 회색빛을 띤다. 1998년에 애리조나주 남동부의 아파치 국유림과 뉴멕시코 남서부의 힐라 국유림에 다시 방사되었다. 현재 야생의 개체수는 35 ~ 50마리로 추정되며, 사육중인 개체수는 200 ~ 300마리로 추산된다. 현재 보호를 받고 있으나 농장주나 목동들로 인해 위기에 처해 있다.
- 툰드라늑대 Canis lupus albus : 털은 검은색, 은회색, 회색이다.
- 늑대 Canis lupus chanco : 몽골, 중국, 러시아 동부에 서식한다. 한반도에선 일제 강점기와 한국전쟁 등을 거치며 멸종한 것으로 추정된다.
독립종
- 동캐나다늑대(Canis lycaon)
재정리 논의 중인 아종
멸종한 아종
가축화

실제 늑대를 길들이기 시작한 과정은 논쟁 중이다. 일반적으로 개는 품종개량의 결과로 생각되나, 늑대 성체가 일반적으로 사람이 길들이기 어려운 것으로 보아서 중석기 시대 인간이 정착지를 이루기 시작했을 때 늑대가 새로운 생태적 지위(패총이나 쓰레기 매립지를 먹이로 삼는 등)를 이루었다는 자연선택이라는 이론도 있다. 늑대와 인간이 편리 공생을 이루었을 때, 수 세대 동안 늑대는 쓰레기를 먹고 자연 선택으로 인간으로부터의 도주 거리가 짧아지고 큰 먹이 사냥을 위한 중복된 기능이 사라지며 신체 변화를 일으켰다는 이론이다.
개는 늑대의 가장 가까운 친척 관계(회색늑대와 개 사이의 유전적 분산은 1.8%이며 회색늑대와 에티오피아늑대, 코요테 사이는 4%이다)이며, 개와 늑대의 행동 및 해부학적 구조 연구에서 개의 생리 및 유전성숙 및 아형보유 구조가 대부분 늑대와 비슷하다. 늑대의 청각융기는 볼록하며 원형이지만, 개의 청각융기는 작고 압축되었으며 약간 뒤틀렸다. 같은 개와 늑대의 크기를 비교할 시, 늑대는 20% 더 큰 두개골과 10% 더 큰 뇌 크기를 가졌다. 이 이해 부위의 노출은 뇌의 중간 부분에 자리를 잡았다. 늑대의 이빨도 개와 비례해서 크며, 소구치와 대구치는 늑대가 덜 빽빽하고 더 복잡한 교두 패턴을 이룬다. 개는 전미골부샘(pre-caudal gland) 기능이 부족하고, 개가 대부분 1년에 두 번 발정하는 데 비해 늑대는 1번 발정한다.
혼종

개와의 혼종
개와 늑대는 유전적으로 가까우며, 수천 년 동안 많은 부분을 같이 생활해왔으나 보통 두 종은 일반적으로 야생에서 교배되지 않았다. 그러나 외톨이 늑대가 경비견이나 양치기 개가 다른 늑대 대신 무리를 이루는 경우가 있다. 그들은 이후 세대에서 풍부한 자손을 만들 수 있었다. 만년에서 15,000년 전 북아메리카에서 늑대와 개의 혼혈로 흑색늑대가 이루어졌다는 것이 밝혀졌다. 유럽의 늑대-개 혼혈화는 늑대의 혈통의 위협성을 느낀 늑대 보호 단체가 우려를 제기했으나, 일련의 미토콘드리아 DNA 분석에서 개의 유럽늑대 침투교잡은 혈통에 상당한 위협이 되지 않는다는 연구가 나왔다. 또한, 늑대와 개의 발정기는 완전히 일치하지 않고 야생 늑대와 개의 혼혈 자손은 나올 가능성이 매우 적다는 것이 증명되었다. 순수 늑대처럼 혼혈종도 매년 번식하고 그 종의 짝짓기 계절은 석 달 전에 일어나나 주로 새끼는 겨울에 태어나기 때문에 이는 그 종의 생존 가능성을 줄게 하였다.
또한, 일반적으로 이누이트 부족들이 늑대의 체력을 개선하기 위해 늑대와 썰매개를 교미시켰다는 주장이 있지만, 이는 사실이 아니고 늑대의 혼혈화는 늑대의 체력이 썰매개보다 약하기 때문에 썰매 등 당기는 장비에 효과적으로 협조할 수 없다. 미국에서 확산한 늑대와 개의 포획 사육으로 약 30만 마리의 늑대개가 탄생했다. 대부분 이러한 목적으로 사용되는 개 품종은 스피츠 종이다. 적어도 두 종의 늑대개 품종인 자르늑대개(Saarlooswolfhond)와 체코슬로바키아늑대개(Czechoslovakian Wolfdog)는 유럽에서 모두 독일산 셰퍼드와 늑대의 혼혈 해서 탄생했다는 것이 증명되었다.

코요테와 황금자칼과의 혼종
수많은 유전자 연구로 북미의 회색늑대는 코요테와 혼혈화되었다는 것이 밝혀졌다. 늑대의 미소부수체와 미토콘드리아 DNA 연구에서, 미국 북동부와 캐나다 남서부의 광범위한 늑대 종들이 코요테와 혼혈화되었음이 밝혀졌고, 코요테와의 하플로타입은 동쪽으로 갈수록 증가하여 미네소타주에서 50%, 퀘벡주에서 100%를 기록했다. 이러한 혼혈종은 늑대보다 주로 작은 영역을 활보하지만, 코요테보다 몸집도 크고 활동 영역도 크다. 2011년, 48,000마리의 동부늑대(주로 앨곤퀸 주립 공원에 서식하는 종)와 붉은늑대(노스캐롤라이나주에 서식하는 종), 코요테 유전자의 단일염기 다형성 검사 결과, 이들은 회색늑대와는 별개의 종이며 늑대-코요테의 혼혈종이라는 것이 밝혀졌다. 동부늑대가 코요테-늑대 혼혈이 된 시기는 콜럼버스 이전 아메리카 시대 600년에서 900년 전 시작되었다고 추측한다. 동부 늑대는 이후 광범위한 선조인 회색늑대와의 여교잡이 진행되었다. 늑대와 황금자칼과의 혼혈화는 자칼의 미토콘드리아 DNA 연구로 세네갈에서 교잡되었음을 밝혀냈다.
계통 분류
다음은 개과의 계통 분류이다.
| 개과 |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||





























신체학적 특징

해부학

회색늑대는 가느다라면서 크고 깊게 내려가는 흉곽과 경사진 등을 가지고 있다. 복부와 목의 근육이 발달하였다. 팔다리가 길고 강하며, 발은 비교적 작다. 앞발은 5개의 발가락이 있고, 또 각각 4개의 발톱이 있다. 앞다리는 겉보기에 가슴에 눌린 것처럼 보이고, 앞발꿈치는 안으로 굽고 발은 밖으로 굽으며 이 때문에 앞다리와 뒷다리가 모두 같은 줄에서 움직일 수 있게 되었다. 늑대의 발은 다른 갯과 동물의 발보다 더 길다. 이는 신속히 움직일 수 있고 많은 눈이 쌓인 바닥에서 쉽게 움직일 수 있게 한다. 암컷은 수컷보다 얼굴과 이마가 좁고 얇은 목, 약간 짧은 다리, 작은 크기의 어깨를 가지고 있다. 사촌 동물(코요테나 황금자칼)과 비교하면 회색늑대는 넓은 코, 짧은 귀, 짧은 몸통과 긴 꼬리, 체중이 더 많은 특징이 있다.
회색늑대의 머리는 넓은 이마, 강한 턱, 길고 무딘 코를 가지고 있다. 귀는 상대적으로 작은 삼각형 모양이다. 이빨은 크고 강하며, 다른 개과 동물보다 뼈 분쇄에 적합하게 생겼으나 하이에나 보다는 약한 힘을 가지고 있다.송곳니는 강하면서 상대적으로 짧다(26mm). 늑대의 이빨은 750psi의 저먼 셰퍼드보다 강한 1,500psi의 압력을 가할 수 있다. 이 힘은 뼈 대부분을 부러뜨릴 수 있다. 추운 기후에서, 늑대는 체온을 보존하기 위해 피부 근처의 혈액량을 줄일 수 있다. 발바닥(footpads)의 체온은 나머지 몸의 체온과 달리 조절할 수 있으며, 이는 얼음이나 눈과 접촉하면서 발바닥이 동상에 걸리지 않게 한다.
회색늑대는 보통 다른 동물에게 경고할 시에만 머리를 높게 든다. 보통은 느린 속도로 이동하나, 다른 생물체가 있을 때에는 발 하나를 더 이용한다. 이 걸음은 시간당 8–9 km/h의 속도를 유지할 수 있고, 이 때문에 늑대는 먼 거리를 이동할 수 있다. 맨 길에서는 늑대가 50–60 km/h의 속도로 달릴 수 있다. 뛰는 늑대는 머리를 한쪽으로 젖혀지고 낮게 하며, 한쪽 귀를 앞, 뒤로 움직이게 한다. 이 자세는 늑대가 지속해서 뛰어난 청각을 활용할 수 있게 한다.
크기

회색늑대는 특정 대형견을 제외하고 개과 동물 중 크기가 가장 크다. 회색늑대의 체중과 크기는 세계적으로 다르나 베르그만의 법칙에 따라 위도와 체중은 비례하고, 이에 따라 알래스카와 캐나다의 늑대는 중동과 남아시아의 늑대보다 3–6배 더 무겁다. 평균적으로 늑대 성체의 높이는 105–160cm이며 어깨높이는 80–85cm이다. 꼬리의 길이는 몸통과 머리 길이의 ⅔로 29–50 cm 정도의 길이이다. 귀의 길이는 90-110mm이며, 뒷다리의 길이는 220–250mm이다. 두개골의 길이는 평균 22~27cm이며, 폭은 12~15cm이다. 늑대의 체중은 지역별도 다양하나, 평균적으로 유럽 늑대는 38.5 kg, 북아메리카 늑대는 45 kg, 인도와 아랍 늑대는 25kg이다. 암컷 늑대들은 수컷 늑대보다 체중이 2~4 kg 더 적다.
드물게 알래스카, 캐나다, 소련 이후 국가들에서 54 kg 이상의 초대형 늑대들이 발견된 적도 있다. 북아메리카에서 가장 무거운 늑대는 1939년 7월 12일 중앙 알래스카에서 사냥 된 회색늑대로 79.4kg이었고, 유라시아에서 발견된 가장 무거운 늑대는 제2차 세계 대전 이후 우크라이나 SSR의 폴타바주 코벨랴키(Kobeliaky)에서 사냥 된 늑대로 약 86kg이었다.
털

회색늑대는 조밀하고 보송보송하며, 짧고 긴 잔털을 가지고 있다. 잔털과 일부 긴 털은 봄에 털갈이하고 가을에 다시 자란다. 긴 털이 목의 앞부분 일부와 목의 뒷부분에 많이 자란다. 특히 목 위쪽과 어깨에 긴 털이 생겨 문양을 만든다. 뺨에는 긴 다발의 털이 있다. 귀는 짧은 털로 골고루 덮여 있다. 팔꿈치 밑 아킬레스건에는 짧고 조밀한 털로 덮여 있다. 북부 지역의 기후에서 늑대가 뒷다리 사이 얼굴을 끼워 넣고 꼬리로 얼굴을 덮을 경우 −40° 온도에서 버틸 정도로 겨울의 털은 추위에 매우 강하다. 늑대 털은 개의 털보다 단열성이 높으며, 따뜻한 숨이 응축될 때 표면에 붙으면서 얼음이 되지 않는다. 따뜻한 기후에서, 털은 북부늑대보다 더 드물게 자란다. 암컷 늑대는 수컷 늑대보다 다리가 부드러운 털로 덮여 있고, 일반적으로 매끄러운 털을 가지고 있다. 늙은 늑대는 코에서부터 이마를 따라 흰 털이 나 있다. 겨울 털은 암컷이 배 주변의 털이 짧음에도 불구하고 평균적으로는 길다. 머리 한가운데의 털 길이는 60–70mm이다. 어깨를 보호하는 털의 길이는 일반적으로 90mm를 넘지 않지만, 일부는 110–130mm도 도달할 수 있다.
털의 색은 다양한 명암이 있어 순수한 하양부터 크림색, 회색, 갈색, 검정까지 다양한 색이 있다. 털 색의 변화는 위도가 높아질수록 증가하는 경향이 있다. 암수 차이에 따른 털 색의 차이는 거의 없고, 암컷의 털이 약간의 붉은 색조가 있다. 털 색은 카무플라주의 목적이 아니며, 일부에서는 늑대 털의 혼합된 색상들이 상호 작용하여 특정 동작을 강조한다는 주장도 있다.흑색늑대(늑대개 혼종 중 하나)는 개와 늑대의 상호 작용으로 인한 야생 늑대 개체 수의 감소로 유라시아에서는 거의 관찰되지 않는다. 흑색늑대는 북미에서 더욱 일반적인 표본이 되며, 옐로스톤 주립공원의 늑대 개체 수 절반 이상이 흑색늑대이다.
감각 능력
회색늑대의 후각은 일부 사냥개에 비해 약하지만, 맞바람이 부는 2–3km에서 떨어진 고기 냄새를 맡을 수 있다. 이 때문에 토끼나 새의 숨겨진 자취를 찾을 수 있다. 청각은 여우보다 더욱 선명하게 듣고, 26kHz의 주파수까지 들을 수 있으며 이는 가을의 잎이 떨어지는 소리도 들을 수 있다.도시전설에서 늑대가 현악기의 소리를 무서워한다는 것은 어느 정도 근거가 있으며, 사육 늑대의 경우 낮은 소리의 현악기 화음을 듣고 고통을 느낀다는 결과가 있다.야간 시력은 다른 개과 동물보다 더욱 뛰어나다.
행동
사회적, 지리적 행동

비록 몇몇 혼자 생활하는 늑대도 존재하지만, 늑대 대부분은 사교적인 동물이다. 늑대의 기본 사회 단위는 한 쌍의 성인 자손으로 구성된 무리이다. 이상적인 조건에서, 서로 만난 한 쌍은 매년 새끼를 낳으며 새끼가 성체가 되어 흩어지기 전 10–54달 동안 무리에 속하게 된다. 평균적인 무리의 동물들은 성체 1–2마리와 청소년 3–6마리, 새끼 1–3마리로 구성되어 5–11마리 정도 이루며, 일부 2, 3개 정도의 무리는 최대 42마리의 늑대로 구성된 거대 무리도 있다. 새끼들이 자라면서 무리 내에서의 성성숙과 경쟁의 시작으로 무리에서 흩어지기도 한다. 새로운 무리는 보통 서로 무관한 암컷과 수컷이 만나 이루어지며, 다른 적대적 무리가 없는 곳으로 떠난다. 늑대 무리는 거의 다른 늑대들을 감싸지 않고 대부분 죽인다. 다른 늑대를 맞이하는 드문 경우에는, 거의 미숙한 동물(1–3년생 정도)이며 쌍들이 사육 권리를 가지기 위해 서로 경쟁한다. 어떤 경우에는, 떠돌이 늑대들이 다른 늑대의 사망 후 무리에 들어가기도 한다. 우제류가 풍부한 시기 동안(이주, 분만 등), 다른 늑대 무리의 늑대가 한때 무리에 들어갈 수 있다.
늑대는 매우 넓은 영역을 가지고 있는 동물이며, 생존을 위해 안정적인 먹이의 공급이 필요한 최소 지역보다 더욱 큰 지역을 가진다. 이 지역의 크기는 사냥 가능한 먹이의 양, 무리 새끼의 나이에 따라 달라지며 먹잇감이 적은 경우나 새끼가 6개월이 되어 성인과 비슷한 영양 요구량을 가질 때 지역의 크기가 커지게 된다. 늑대 무리는 먹이를 찾아 계속 이동하며, 이는 하루 영역 25 km/d의 9% 정도를 차지한다. 늑대의 삶 전체 동안 이용하는 핵심적인 평균 영역은 35Km²이며, 평균 50%를 이곳에서 보낸다. 먹이 밀도는 이 지역의 주변 지역에서 훨씬 높은 경향이 있으며, 늑대는 가능한 한 지역의 가장자리에서의 사냥은 다른 늑대 무리와의 심각한 조우가 있을 가능성이 높아 피하는 경향이 있다. 기록상 가장 작은 영역은 미네소타 북동부의 6마리 늑대들로 구성된 무리로 33Km²의 영역이며, 가장 큰 영역은 알래스카의 10마리 늑대로 구성된 무리로 6,272Km²이다. 일반적으로, 늑대 무리는 자기 지역에서 정착하게 되며 심각한 식량 부족 사태가 발생할 때까지 계속 익숙한 범위에서 맴돌게 된다.
늑대는 영역 표시를 하며 울부짖거나 직접 공격으로 자기 영역을 지킨다(늑대#의사소통을 참조하라.) 늑대는 자기의 냄새와 채취로 영역을 표시한다. 늑대가 묻힌 채취는 자기 영역에 걸쳐 240m 밖에서도 느낄 수 있다. 이러한 채취는 2–3주 동안 지속되고, 보통 바위나 나무, 큰 동물 뼈에 묻힌다. 채취 표시나 하울링 등이 실패할 경우, 다른 늑대 무리로부터 지역을 지키기 위해 서로 싸움을 하기도 한다. 미네소타 주와 데날리 국립공원에서 사망한 늑대 중 14–65%가 다른 늑대와 싸운 것이 원인일 정도로 싸움은 늑대 사망의 주요 원인이다.
생식 및 출산

일반적으로, 회색늑대는 단혼이며 쌍의 둘 중 한 마리가 사망하지 않는 이상 계속 같이 살아간다. 한 쌍의 늑대 중 한 마리가 사망하면 곧바로 다른 쌍을 찾는다. 종종 수컷은 특정 늑대에서 우위를 차지하기에, 짝이 없는 암컷은 드물다. 만약 수컷 늑대가 자기 영역에서 짝을 찾지 못할 경우, 이 늑대는 다른 늑대 무리에서 번식한 암컷 늑대와 짝짓기한다. 이러한 늑대를 "카사노바 늑대"라고 하며, 늑대 무리의 수컷과는 달리 이들은 암컷과 교미하여 페어 본드를 만들지 않는다. 몇몇 옐로스톤 국립공원의 늑대 무리는 암컷이 여러 번 번식하기도 한다.
회색늑대에게도 동성애 습성이 관측된다. 무리 내에서 가장 지위가 높은 암컷에게 발정기가 찾아오면 수컷 늑대가 다른 수컷 위에 자주 올라타곤 한다.

늑대의 첫 번식 시기는 환경에 따라 달라지나, 먹이가 풍부하거나 늑대 개체 수가 많을 경우 늑대는 더욱 풍부한 자원을 이용하기 위해 어린 시기에 번식하기도 한다. 이것은 사육 늑대가 9–10개월 이내에 번식 시기에 도달하고, 야생의 최연소 번식 늑대는 2년이라는 것으로 알 수 있다. 암컷은 매년 생식이 가능하나 매년 평균적인 개수의 동복자를 만든다. 코요테와는 달리, 회색늑대는 결코 폐경기가 찾아오지 않는다. 일반적으로 발정은 겨울에 나타나고 출산 경험이 있는 암컷은 2–3주 이전 정도 젊은 암컷보다 빨리 찾아온다. 임신 중, 암컷 늑대는 다른 무리와의 폭력적인 조우를 피하려고 자신 영역의 가장자리로 이동하지 않는다. 늙은 암컷은 보통 동복자의 동굴에서 새끼를 나고, 젊은 암컷은 자신의 동굴에서 출산한다. 임신 기간은 62–75일이며, 새끼는 보통 여름에 출산한다.
늑대는 다른 개과 종과 비교해 작은 동복자에서 많은 새끼를 출생한다. 평균적으로, 하나의 동복자에 새끼 5~6마리가 있고, 먹이가 풍부할 경우에는 돌보는 새끼가 많아지며 동복자도 증가하지만, 14–17마리 정도의 매우 많은 새끼는 1% 이하에 불구 하다. 새끼는 보통 봄에 태어나 먹이에 상응하는 인구 증가세를 보여준다. 새끼는 태어날 땐 실명과 청각 장애 상태이며 짧고 부드러운 진갈색 털로 덮여 있다. 출생 당시 몸무게는 300–500g이며, 9–12일 후 앞을 보기 시작한다. 송곳니는 한 달 후 나타나기 시작한다. 새끼는 처음 3주 동안 동굴에 둔다. 1달 반 후, 새끼는 위험으로부터 도망갈 수 있을 만큼 민첩해진다. 어미 늑대는 처음 몇 주 동안 동굴을 떠나지 않고 젊은 늑대가 주는 음식에 의존한다. 새끼는 3–4주 후부터 딱딱한 음식을 먹기 시작한다. 새끼는 출산 후 첫 4달 동안 무게가 30배 증가하며 매우 빠른 성장 속도를 보인다. 늑대 새끼는 3주부터 싸우는 연습을 시작하지만, 이와 달리 여우나 코요테는 최대한 무는 것을 줄인다. 일반적으로, 실제적인 싸움은 위아래 계층 구조가 만들어지기 시작하는 5–8주 후부터 시작된다. 코요테나 여우와는 달리, 늑대는 놀이 행동이 나타나기 전부터 싸움 행동이 나타나기도 한다. 가을이면, 새끼는 큰 먹이를 사냥하며 성체와 같이 사냥에 동반 가능할 정도로 성장한다.
굴과 주거지의 행동
늑대는 굴에서의 휴식은 춥거나, 습하고 바람이 강한 날씨일 때 쉬고 낮에 건조하거나, 조용하고 따뜻한 날씨에 늑대는 공개적인 장소에서 휴식한다. 봄가을 동안 늑대는 더욱 활발해지며 자신의 영역 아무 곳에서나 공개적인 곳에서 눕기도 한다. 실제로, 굴은 여름 동안 새끼를 키우기 위해 만들어졌다. 굴을 구축할 때 암컷은 바위의 갈라진 곳 등 자연적으로 만들어진 곳에 굴을 만들고 절벽이나 돌출된 구멍을 두꺼운 식물로 가린다. 때때로, 이 굴은 여우, 오소리나 마멋 등 작은 동물이 서식하기에 적합한 굴이기도 한다. 이 적합한 굴은 종종 확장하거나 부분적으로 고치기도 한다. 드문 경우지만, 암컷이 스스로 작은 1–3개의 굴을 파기도 한다. 늑대는 기생충 예방 차원에서 여러 개의 굴을 같은 위치에 만들지 않는다. 굴은 보통 물에서부터 500m 떨어져 있는 곳에서 만들고, 보통 남쪽으로 굴을 만들어 햇빛이 잘 들어오게 하며 상대적으로 쌓인 눈들을 자유롭게 두기도 한다. 휴식 장소, 새끼를 기르는 곳, 먹이 보관 굴 등은 일반적으로 늑대 굴 주변에다 둔다. 굴 주변에서 나오는 악취 등은 까치나 큰까마귀 등의 조류를 유도하게끔 한다. 굴을 위해 필요한 곳이 주변에 있어 늑대 굴에는 보통 가족 동물도 같이 산다. 늑대는 주로 인간의 눈을 피해 굴을 짓지만, 늑대는 건축물, 도로, 철도 주변에 굴을 짓는 것으로 알려졌다.
지능

비록, 늑대의 지능 연구는 정량화되지 않고 흩어져 있는 연구와 소규모 실험실 연구가 있지만, 종의 기억 능력과 연관 사건을 기억한다는 연구가 있다.

그레이트플레인스에서 아메리카들소가 멸종되는 동안, 늑대는 총소리를 따라 하는 법을 익히고 사냥꾼이 먹이를 가져가기 전에 먼저 들소를 가져가게 되었다. 북부 몬태나 주에서, 늑대가 공중의 사냥꾼들에게 공격받자 늑대들은 항공기 소리를 들을 때마다 개방된 장소에서 도망치게 되었다. 반대로, 항공기를 이용한 연구에서 늑대가 공격하지 않을 것이란 확신을 얻을 경우에는 매우 낮게 근접하는 경우에도 두려워하지 않는 것을 발견했다. 사냥할 때, 무스나 작은 사슴 두 종이 있으면 사냥하기 쉬운 후자를 사냥하는 경우가 더욱 많다. 늑대의 기억력은 동물학자 조지 미바르트가 길들어진 늑대에 대해 3년 동안 떨어져 있었던 후 다시 만날 때도 주인을 알아본다는 것이 발견되었다. 과격하지 않은 늑대에 대해, 늑대는 무장인과 비무장인을 구별할 수 있으며, 전형적인 무장인인 경우의 태도에 확신하지 않을 때에는 그 사람들을 공격하지 않는다. 성체 늑대는 굴을 만들 때 숲이나, 바람, 썩은 나무, 기타 자연물 등을 이용하여 능숙하게 숨기는 법을 알고 있는 것으로 추측된다. 함정에 걸린 적이 있던 늑대의 경우, 함정을 피하거나 피해 없이 함정을 건드리기도 한다. 예를 들어, 커럼포우의 왕 로보 이야기(Lobo the King of Currumpaw)에서 자연주의자 어니스트 톰프슨 시턴은 초기 늑대를 잡는 시도가 실패할 경우 늑대는 함정을 피하거나 무시한다고 적었다.
늑대는 훈련할 수 있지만, 개에게서 보이는 만큼의 훈련도를 얻기는 힘들다. 늑대는 불쾌한 자극과 힘에 대해 민감하게 반응하고 두려워하지 않는다. 일반적으로, 개 대부분에게서 얻는 신뢰도보다 더 많거나 비슷한 일을 해야 한다. 심지어는, 늑대는 특정 명령을 반복해서 받는 경우 지루해하며 명령을 듣지 않을 수 있다. 늑대는 긍정적인 반응과 보상에 더욱 많은 반응을 보이지만, 간단한 칭찬 같은 경우에는 다른 개과 동물과 달리 충분하지 않게 받아들인다. 개와 달리, 늑대는 음성보다 수신호에 더욱 반응하는 경향이 있다. 늑대를 훈련하는 대부분의 시도는 작업견 등에서와 같은 실패를 불러왔다. 독일 생물학자 에릭 지먼은 자신의 사육늑대를 썰매개로 만들고자 시도했고, 늑대는 마구를 매고 적극적으로 썰매를 끌긴 했으나 개인 공간의 확보 위해 서로 싸우고 한 번 지치면 명령을 듣지 않으며 다른 야생 동물에 의해 집중도가 떨어졌기 때문에 궁극적으로는 신뢰할 수 없었다. 그러나 존 제임스 오듀본은 켄터키주에서 늑대에게 사슴 사냥 훈련이 가능했다는 기록을 남겼고,헨리 와튼 슈메이커(Henry Wharton Shoemaker)는 서부와 중부 펜실베이니아주의 정착민들은 늑대를 사냥개로 이용했다는 기록이 남아 있다.조르주루이 르클레르 드 뷔퐁이 쓴 "자연사"에서, 페르시아의 늑대들은 춤을 추도록 훈련된다는 기록을 남겼다.
사냥 및 먹이 행동


비록, 늑대는 사회적 동물이지만 큰 무리보다 두 정합 쌍의 무리가 더욱 사냥 확률이 높고, 혼자 있는 늑대의 경우에는 들소나 사향소 같은 자기보다 큰 먹이를 죽이는 경우도 있음을 밝혔다. 늑대 사냥은 다음의 5단계로 나눌 수 있다.
- 먹이 탐색: 늑대는 냄새로 먹이를 찾기 위해 돌아다니고 추적한다. 일반적으로, 늑대는 냄새로 먹이를 찾으며 직접 바람이 불어오는 쪽의 먹이 냄새만 맞을 수 있다. 먹이의 냄새가 바람과 일치할 경우, 늑대는 서서 목표를 향해 눈, 귀, 코를 그 위치로 가리킨다. 열린 지역에서, 늑대는 무리를 지어 코와 코끼리 서 있고 꼬리를 흔들며 앞에 선다. 일단 늑대들은 먹이를 향해 달려가는 것으로 결론을 내린다.
- 몰래 접근: 늑대는 자신을 은폐하면서 먹이에 다가간다. 늑대와 먹이 사이의 간격이 좁히기 위해 꼬리를 흔들며 빠르게 뛰며, 먹이를 도망치지 못하게 하도록 자신의 포위망으로 움직이게 한다.
- 맞닿드임: 먹이가 늑대를 감지하게 되면, 도망 또는 늑대에게 접근하는 두 가지 입장이 있다. 늑대보다 더 큰 몸집의 엘크나 무스, 들소 같은 경우에는 그 자리에서 가만히 서 있다. 문제가 생길 경우, 늑대는 돌아가며 먹이가 도망갈 경우 등 자극을 줄 경우에는 공격을 시작한다. 만약 동물이 그 자리에서 설 경우에는, 늑대는 이를 무시하거나 위협하게 된다.
- 돌진: 먹이가 도망 갈려 할 경우, 늑대는 즉시 따라가며 공격한다. 이 단계는 늑대가 계속 최고 속도로 달릴 수 없으므로 사냥에서 제일 중요하다. 먹잇감이 단체로 있는 경우, 늑대들은 이 무리를 떨어뜨리거나 한두 마리의 먹이들을 분리한다.
- 추적: 계속 돌진한 후 따라잡힌 경우에는 늑대가 먹이를 죽이려고 한다. 작은 먹이를 쫓는 경우, 늑대는 가능한 한 빨리 먹이를 잡으려 하며 큰 먹이를 쫓는 경우에는 추적이 길어지고 선택한 먹이에 대한 순서를 유지한다. 보통, 늑대는 1–2 km 정도 추적한 후 포기하지만 한 늑대의 경우에는 사슴을 21km나 추적한 후 잡은 적도 있었다. 러시아와 북아메리카의 늑대 모두 먹이의 속도를 늦출 수 있는 얼음, 절벽, 계곡, 가파른 경사 지역으로 모는 것이 관찰되었다.
옐로스톤에 늑대가 처음 재도입되었을 때 처음에는 엘크사슴만을 사냥했었다. 재도입된 늑대무리들끼리 서로 싸움이 일어나면서 그 때 당시 큰 세력을 형성했던 드루이드 봉우리 무리가 몰리 무리와 다른 무리를 몰아내면서 엘크사슴이 풍부한 라마 계곡(Lamar Valley)을 독차지하게 되었고, 몰리 무리를 비롯한 다른 무리는 엘크사슴이 풍부한 곳을 빼앗기고 들소가 흔한 지역으로 쫓겨나면서 들소를 사냥하게 되었다. 처음에는 들소 사냥에 서툴렀지만, 점차 들소 사냥을 하는 방법을 쌓게 되었으며 현재는 진흙탕 샛강 무리(Slough Creek Pack)가 대표적인 들소 사냥꾼이다. 북극 지역의 경우에는 들소나 엘크사슴 보다는 순록, 사향소, 레밍쥐를 비롯한 설치류와 토끼를 흔히 사냥한다. 철마다 오는 순록으로 늑대들은 늙거나 병든 개체를 사냥함으로써 순록과 사람(크릭 족이나 내륙의 이누이트들 같은 북극권 수렵 원주민들)에게 건강한 개체를 남겨 줌으로써 이득을 가져다준다. 순록이 없을 때에는 주로 설치류를 잡아먹는데 순록사냥이나 사향소 사냥처럼 무리를 짓지 않고 홀로 찾아서 먹는다. 새끼를 키울 때도 쥐고기로 키운다. 늑대들은 어류도 잡아먹는다. 한 예로, 알래스카 남서부 끝에 있는 큰곰자리 숲이라는 곳에서 늑대들은 연어들이 산란하러 올 때마다 연어를 사냥한다. 송어 또한 이들의 먹잇감이다. 이것은 그들의 덩치 큰 친척인 다이어늑대와 대조되는 특징이다. 다이어늑대들은 커다란 들소나 야생마를 주로 잡아먹었기 때문이다. 에티오피아의 아비시니아 고원에 사는 늑대들은 무리를 지어 살지만, 사냥을 할 때에는 독자적으로 나서서 그곳에 흔한 설치류들을 사냥한다.
실제로 죽이는 방법은 종마다 다르다. 몸집이 큰 먹이일 경우, 성체 늑대는 정면 공격을 피하고 동물의 측면이나 후면을 노린다. 순록 같은 대형 먹이는 부드러운 회음 부분을 물려 과다 출혈로 사망한다. 이러한 상처의 길이는 보통 10~15cm이며, 건강한 사슴도 쓰러질 정도로 출혈이 심해지는 회음 부분을 문다.양이나 노루 같은 비슷한 크기의 먹이를 잡을 경우, 늑대는 신경계와 목동맥을 물어 몇 초 만에 먹이를 죽인다. 쥐와 같은 작은 먹이를 사냥할 경우, 높은 곳에서 재빨리 뛴 다음 앞발로 고정해 움직이지 못하게 잡는다. 먹이들이 사냥하기 쉽고 풍부한 경우에 늑대는 가끔 과잉 살해를 하기도 한다. 이러한 경우는 개에게도 많이 찾아볼 수 있지만, 야생의 경우에는 매우 드문 일이다. 야생에서, 과잉 살해는 보통 늦겨울이나 봄에 눈이 유난히 많이 쌓인 경우(그러므로 먹이가 잘 움직이지 못함) 또는 굴에서 양육하는 기간 늑대가 많은 수의 먹이가 있어야 하는 경우 일어난다. 늑대가 목을 물어 빨리 죽일 수 있는 중간 크기의 먹이들이 과잉 살해에 가장 취약하다. 성체 늑대가 새끼에게 사냥을 가르칠 때에도 과잉 살해가 발생한다.
일단 먹이가 쓰러지면, 늑대는 흥분하여 시체의 모든 방향에서 달려들어 먹이의 큰 덩어리 부분부터 먹기 시작한다. 번식 중인 쌍의 늑대는 새끼를 계속 낳기 위해 음식을 독점한다. 먹이가 부족하면, 이는 늑대 구성원 중 새끼에게 가장 큰 부담이 된다. 일반적으로 먹이를 잡은 번식 중인 쌍이 먼저 먹이를 먹은 이후에는 오랜 사냥 후 나머지 구성원들이 방해받지 않고 먹을 수 있게 한다. 번식 중인 쌍 늑대가 먹은 후에는, 나머지 구성원들이 시체 조각들을 찢은 다음 한적한 지역으로 옮긴다. 늑대는 심장, 간, 허파, 위 내벽 등 먹이의 큰 내부 기관부터 먹기 시작한다. 콩팥과 비장은 밖으로 노출되면 먹고, 그다음에는 근육을 먹는다. 한 마리의 늑대는 한 번에 자기 몸무게의 15–19% 정도의 먹이를 섭취한다.
의사 소통
시각적
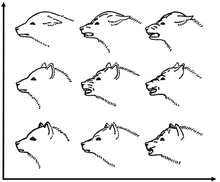
회색늑대의 의사소통 행위는 단체 생활과 사냥 습관 때문에 코요테나 황금자칼보다 복잡하다. 덜 사교적인 개과 동물은 간단한 시각적 신호로 의사소통하지만, 늑대는 더욱 자세하고 미묘한 시각적 신호가 있다. 자연적일 때, 늑대는 다리를 굳히지 않고 꼬리를 느슨하게 내리며 얼굴이 부드러워지고 귀는 특별하지 않은 방향으로 가리킨 채로 서 있다. 늑대의 행동 의사소통은 꼬리와 소름 등의 다양한 구성으로 표현한다. 공격적이거나 자기주장이 강한 늑대는 움직임이 신중하고 느린 것이 특징이며, 몸이 높은 자세이며 목뒤털이 올라가지만, 복종하는 늑대인 경우에는 자세가 낮고 자신의 털, 귀, 꼬리를 낮춘다. 번식 가능한 수컷은 자기 무리 부하의 가족을 보았을 때 꼬리를 척추와 수평으로 든다. 복종하는 행동은 수동적 과 능동적 두 형태로 이루어진다. 수동적인 방식은 보통 지배적인 동물이 다가왔을 때 보여주는 방식이며, 복종하는 늑대는 지배적인 늑대가 회음의 냄새를 맡도록 한다. 능동적인 방식은 지배적인 늑대를 만났을 때 낮은 자세로 접근하며 다른 늑대의 얼굴을 핥는 식으로 인사하는 방식이다. 늑대가 함께 있을 때, 일반적으로 이들은 코나 턱을 서로 밀거나 뺨을 맞대고 얼굴을 핥는 행동 등을 한다. 서로의 코 부분에 입을 대는 것은 친절을 표시하는 것이고, 이빨이 코 부분에 닿는 것은 지배를 뜻한다.
| 신체 부위 | 공격적 | 공포 |
|---|---|---|
| 눈 | 휘둥그레진대로 직접 쳐다봄 | 눈을 길게 하여 먼 곳을 쳐다봄 |
| 귀 | 똑바르고 앞으로 섬 | 평평하고 펴저 있음 |
| 입술 | 가로로 수축됨 ("무리한 주름"(agonistic pucker)) |
수평으로 펴짐 ("복종의 미소"(submissive grin)) |
| 입 | 열림 | 닫힘 |
| 이빨 | 송곳니 노출 | 송곳니를 덮음 |
| 혀 | 뒤로 말림 | 펼쳐짐 ("핥기 위한 의도") |
| 코 | 짧아짐 (피부가 접힘) | 길어짐 (피부가 늘어남) |
| 이마 | 수축함 (눈쪽이 볼록해짐) | 커짐 (늘어남) |
| 머리 | 높아짐 | 낮아짐 |
| 목 | 아치 모양 | 길어짐 |
| 털 | 똑바로 섬 (곤두섬) | 매끄러움 |
| 몸 | 높이 섬 | 낮게 낮춤 |
| 꼬리 | 높이 듬 떨림 |
몸 아래로 내림 흔들림 |
청각적

하울링 (Howling)
늑대는 보통 사냥 직후 무리를 다시 모이게 할 때 정보를 알리기 위해, 특히 굴에서 익숙하지 않은 지형에도 서로 의사소통을 하게 할 수 있도록 소리를 이용한다. 늑대의 울부짖음은 가장 좋은 조건에서 최대 130 km3 밖의 영역에서도 들을 수 있다. 늑대의 울부짖음은 큰 개의 소리와는 구별된다. 수컷 늑대는 베이스 O 소리로 짖고, 암컷 늑대는 코에서 바리톤 U 소리를 낸다. 새끼는 울부짖지 않으며, 한 살 정도의 어린 늑대는 옙(yelps)같이 개와 비슷한 짧은 울음소리를 낸다. 늑대 울부짖음의 주파수 대역은 150 ~ 780Hz를 기본 주파수로 하여 12음 위의 함축된 화성을 낸다. 보통 피치는 일정히 유지되나, 4~5배 정도 높게 올리거나 내릴 수 있다. 이 울부짖음은 무리를 부를 때 사용하며, 부드러운 소리인 시작되는 소리는 수리부엉이의 소리와 비슷하다. 먹이를 쫓는 중에는 고음에서 두 음을 오가는 울부짖음을 낸다. 먹이가 가까워지면 짧은 짖음과 울음소리를 혼합하여 낸다. 함께 울부짖을 때에는 늑대가 같은 수에서 조화롭게 합창하여 실제보다 더 많은 늑대가 있는 것처럼 속인다. 외톨이 늑대는 일반적으로 다른 늑대 무리가 있는 지역에서는 울부짖지 않는다. 늑대들은 비가 오는 날씨거나 자기가 만족스러운 경우에는 울부짖음에 대답하지 않는다. 유럽 늑대는 북미 늑대보다 훨씬 더 길고 크며, 첫 번째 음절을 강조하는 등 늑대의 울부짖음은 지역마다 다르다는 주장이 있다. 그러나 둘은 상호 의사 소통성으로 북미 늑대가 유럽 늑대의 울부짖음의 차이점을 기록하기 위해 만들어진 것이다.
기타 발성
늑대의 여러 소리에는 으르렁거리는 소리(growls), 짖는 소리(barks), 낑낑거리는 소리(whines) 3가지로 나눌 수 있다. 짖는 소리는 320–904Hz를 기본 주파수로 하여 놀란 늑대가 내는 소리이다. 늑대는 개처럼 크고 길게 짖지는 않지만, 몇 번 짖고 나서 위험으로부터 돌아간다. 으르렁거리는 소리는 380–450Hz를 기본 주파수로 하여 늑대가 먹이 문제에 직면했을 때 내는 소리이다. 일반적으로 새끼에게 으르렁거리는 소리가 난다. 늑대의 울부짖는 소리를 동반하며 높은 음조를 내는 낑낑거리는 소리는 공격을 위해 돌진할 때 내는 소리이다. 낑낑거리는 소리는 이 외에도 불안, 호기심, 탐구, 친밀감과 인사, 새끼에게 먹이를 줄 때 등 다양한 때에 나는 소리이다.
후각적

늑대의 후각은 가장 예민하고 근본적인 의사소통의 역할을 한다. 늑대의 얼굴, 입술, 발가락 뒤에는 아포크린선이 많다. 이 내분비선에서 나오는 냄새는 늑대의 미생물상과 식슥관에 따라 다르며, 각자 고유한 후각적 지문을 가진다. 아포크린과 에크린한선이 섞여, 늑대가 땅을 긁으면 그 곳에 냄새가 배이며 이는 배뇨 후와 번식기에 깨끗하게 하기 위해 한다. 늑대 뒷면 머리카락의 모공은 아포크린선과 피지선이 나오는 장소이다. 일반적으로 뒷면의 피부는 접히기 때문에 이 선에서 나오는 물질로 세균이 서식하는 미기후가 만들어진다. 소름이 일어나는 동안, 뒤면의 머리카락이 나오고 피부의 주름이 펴저 냄새가 나오게 된다.꼬지전분비선은 늑대끼리 싸울 때 꼬리에서 나오는 선으로 꼬리를 높이며 분비하므로 가장 높은 지점에 있는 향선이다. 늑대는 항문낭에 아포크린선과 피지선을 모두 가지고 있다. 이 항문낭 분비물의 구성 요소는 계절과 성별에 따라 다르므로, 분비물은 성별과 생식 상황을 알 수 있게 한다. 포피선의 분비물은 호르몬의 상태 또는 사회적 지위를 알려주며, 조사에서 지배적인 늑대는 부하를 통해서 포피선의 냄새를 알려주는 것이 알여졌다. 번식기동안, 암늑대는 질 분비물로 암컷의 생식 상태를 먼 거리의 수컷도 감지할 수 있게 한다. 배뇨는 늑대의 후각적 의사소통에서 가장 중요한 곳이다. 대부분의 연구자들은 이 곳의 기능이 동물들의 경계를 나타낸다는 것에 동의한다. 늑대의 소변은 늑대가 익숙하지 않은 지형에서 더욱 강하게 나타나며, 또한 침입한 영역에서는 다른 늑대나 개과의 향과 같이 난다. 다리 올림 배뇨(RLU)는 암컷보다 수컷이 더 일반적이며, 이는 늑대가 감지 가능성을 극대화할 목적으로 이루어져 있으며 이 마크는 늑대의 높이도 알려준다. 일반적으로 지배적인 늑대가 RLU를 사용하며, 하위적인 수컷은 어린 선 자세를 이용한다.
생태학
식습관

회색늑대는 대형 유제류를 먹이로 하기 때문에 전문종으로 분류한다. 유라시아에서, 인간이 많은 활동을 하는 구역의 늑대들은 가축이나 쓰레기 먹이에 의존하지만, 러시아나 동부 유럽 산악 지역 등 야생의 늑대들은 무스, 말사슴, 유럽노루, 무플론, 멧돼지 등 대형 유제류를 먹이로 삼는다. 기타 초원의 늑대들은 순록, 유럽들소, 사이가산양, 염소속, 샤모아, 들염소, 다마사슴, 사향노루 등을 먹이로 삼는다. 북미 늑대는 쓰레기나 가축에 의존하는 도시 늑대를 제외하면 대부분 인구 밀도가 낮은 지역에서 적당한 서식지를 갖추고 있다. 북미의 늑대가 선호하는 먹이는 무스, 흰꼬리사슴, 와피티사슴, 뮬사슴, 가지뿔영양, 큰뿔양, 돌산양, 흰바위산양, 아메리카들소, 사향소, 순록 등이 있다. 다만 들소와 무스는 몸집이 크고 힘이세서 늑대도 혼자서는 사냥을 못한다.
늑대는 주로 대형 유제류를 먹지만 다른 먹이도 가리지 않는다. 늑대가 보완하기 위해 먹는 소형 먹이로는 아홉띠아르마딜로, 토끼, 여우, 족제비, 아메리카너구리, 오소리, 얼룩다람쥐, 생쥐, 햄스터, 들쥐, 프레리도그, 우드척, 마멋, 비버 등 설치류 및 기타 식충목이 있다. 또한 물새의 알도 자주 먹는다. 이러한 먹이가 부족한 경우, 늑대는 도마뱀, 뱀, 개구리, 드물게는 두꺼비와 대형 곤충도 또한 먹이로 삼는다. 먹이가 부족한 시기에는 소 도축장 등에서 나온 부육을 먹기도 한다. 거친 겨울 동안 종종 무리에서 약하거나 부상당한 늑대를 공격하거나 죽은 무리의 늑대를 먹기도 하는 것 등 동종포식도 종종 있는 일이다.아스트라한의 늑대 무리는 카스피해에서 카스피물범을 사냥한 적도 있으며 알래스카와 서부 캐나다의 늑대 무리들은 연어를 사냥한 적도 있다.인간은 거의 먹이로 노리지 않지만 가끔 공격하는 사례가 있다.
늑대는 장과, 산딸기, 은방울꽃, 월귤나무 열매, 블루베리, 월귤 등 과일과 채소로 식습관을 보충한다. 가지과, 사과, 배나무속의 과일을 먹기도 한다. 여름에는 멜론을 먹기도 한다. 늑대는 먹이를 2주간 먹지 않아도 근육 활동이 약화되지 않을 만큼 오랜 기간 음식 없이 생존할 수 있다. 잘 자란 늑대는 가을-겨울 기간 동안 피부 아래, 심장 주변, 창자, 신장 및 골수에 지방의 형태로 에너지를 저장한다. 늑대는 소화에 몇 시간만 소요되므로 하루에 몇 번이나 먹이를 먹을 수 있고 대량의 고기를 빠르게 소화시킬 수 있다.
천적과 경쟁


일반적으로 늑대는 다른 개과 동물과 있을 경우 지배적인 위치에 있다. 북아메리카에서 늑대는 겨울에 일반적으로 코요테를 죽인다. 늑대는 코요테의 굴을 파고 먹지도 않고 새끼를 죽인다. 코요테는 늑대를 쫓기는 하지만 코요테가 늑대를 죽였다는 기록은 없다. 유라시아와 아프리카에서 발견된 한 지역에서 늑대와 황금자칼와의 상호 작용에는 후자가 수가 더 적고 늑대가 더 많다. 늑대가 주요한 육식 동물인 너구리는 봄가을에 많이 죽인다. 또한 늑대는 붉은여우, 북극여우, 코사크여우를 부육 쟁탈에서 죽이고, 가끔은 먹기도 한다. 아시아에서 늑대는 승냥이와 경쟁 관계이다.

늑대는 유라시아와 북미에서 큰곰과 만난다. 큰곰은 부육 경쟁에서 늑대에 우위 관계이지만 늑대는 서식지에서 큰곰보다 우위에 있다. 두 종은 서로의 새끼를 죽인다. 늑대가 큰곰 새끼를 먹고, 큰곰도 주요 먹이가 늑대 새끼이다. 미국에서는 아메리카흑곰도 마주친다. 흑곰과 늑대가 만나는 일은 서식지 차이로 인해 큰곰과 만나는 일보다 드물다. 흑곰과 늑대가 만난 것 대부분은 멕시코 북부에서 기록된 것이다. 늑대는 수많은 경우에 흑곰의 둥지를 공격하거나 먹거나 죽이려 하지 않는다고 기록되었다. 큰곰과는 달리, 흑곰과 늑대의 살해 논쟁은 아직도 지속되고 있다. 일반적으로 흑곰과 큰곰과의 조우는 많은 편이지만, 북극곰은 늑대 2무리가 북극곰을 죽였다는 기록이 있으나 만났다는 기록은 거의 없다. 늑대는 아시아흑곰의 새끼를 죽였다는 기록이 있다.
중앙아시아와 이스라엘의 늑대는 부육 쟁탈전으로 줄무늬하이에나와 많이 조우한다. 줄무니하이에나는 늑대와 상호 작용 지역에서 늑대의 시체를 먹는다. 일대일에서는 하이에나가 늑대보다 우위이지만 늑대 무리는 하이에나보다 우위이다.
거대한 크기의 늑대 무리는 중간 크기의 고양이아과보다 수가 적다. 늑대는 로키산맥과 그 인접 산맥에서 퓨마와 부딪힌다. 늑대와 퓨마는 일반적으로 다른 고도에서 사냥하므로 서로 잘 만나지 않는다. 그러나 겨울에는 눈이 쌓인 계곡에서 먹이를 사냥하면서 늑대와 퓨마가 만날 확률이 높아진다. 이들은 상호 작용을 거의 하지 않지만 늑대와 퓨마는 서로를 죽이려 하며 늑대 무리는 때때로 퓨마를 죽이기도 한다. 이들은 마눌들고양이를 사냥하며 눈표범은 위협을 준다. 늑대는 스라소니 개체수를 매우 줄일 수 있다.
다른 인간이나 호랑이만이 늑대에게 심각한 위협을 주는 포식자이다.러시아 극동 같이 호랑이와 늑대의 서식지가 겹치는 경우 두 종은 치열한 경쟁을 한다. 늑대와 호랑이의 상호 작용은 20세기 초까지 시호테알린 산맥의 늑대 개체수는 매우 적었다는 기록으로 알 수 있다. 늑대 개체수는 19세기 후반부터 20세기 초반까지 러시아가 식민지가 되어 호랑이 개체수가 감소했을 때만 증가했다. 시호테알린 산맥 주변에 거주하는 원주민들이 1930년대까지 늑대가 서식한 기억이 없으며 이후부터 보이기 시작했다고 증언했다. 호랑이는 생태계 내에서 거의 의미가 없을 정도로 늑대의 개체수가 줄어들게 하거나 지역 멸종을 시켰다. 늑대는 오직 인간이 호랑이를 경쟁적 배제를 시킴으로써만 살 수 있었다. 오늘날 늑대는 호랑이 거주 지역에서 주로 외톨이 늑대나 소형 무리의 형태로 포위되어 서식하고 있다. 호랑이가 자연사해 죽은 경우 늑대는 가끔 청소 동물로서 나타난다. 늑대가 호랑이를 죽인 경우는 매우 드물며, 자연에서 호랑이를 공격한 경우는 최소 4건 있다. 이 호랑이의 늑대 경쟁 제외는 러시아의 호랑이 사냥꾼이 호랑이를 사냥할 수 있는 근거를 만들었고 이들은 늑대 개체수를 유제류 이하로 만듦으로써 늑대 인구도 통제했다.
서식지 및 보존
회색늑대는 인간과 사자 이후로 세계에서 널리 퍼진 동물 중 하나이며, 북위 15°북아메리카에서 북위 12°인도까지 걸쳐 서식하고 있다. 인간의 가축 공격과 두려움으로 인한 고의적인 개체수 감소로 3번째로 빨리 개체수가 감소한다. 이 종은 서유럽에서 멸종했으며, 멕시코와 미국의 늑대로 멸종중이었다. 현대의 늑대는 황무지와 벽지에 서식한다. 특히 캐나다와 알래스카주 등 미국 북부, 북유럽, 북아시아에 거주하며 북위 75°에서 12°에 걸쳐서 서식하고 있다.
늑대 개체수 감소는 전 서식 장소의 재도입과 재정착, 법적 보호, 토지 사용의 변화와 이촌향도 현상으로 억제되었다. 가축 및 게임 종(game food)과 인간의 경쟁과 인간의 늑대에 대한 두려움, 서식지의 조각화로 늑대는 지속적인 위협을 받고 있다. 이러한 위협에도 불구하고, 늑대의 광범위한 범위의 서식지와 안정적인 개체수는 세계적으로 위협적이지 않다고 판단되어 국제 자연 보호 연맹에서는 "관심 필요"의 등급을 받았다.
유럽
쇠퇴

북유럽에서 늑대의 멸종 노력은 중세부터 조직화되어 1800년대 후반까지 계속되었다. 영국에서 늑대는 입법에 의해 박해당했고 브리튼섬의 마지막 늑대는 16세기 헨리 7세 시기 죽었다. 늑대는 이후 광대한 숲이 있는 스코틀랜드로 옮겼으나 이후 숲들이 사라졌다. 늑대는 이후 1684년까지 브레마 주와 서덜랜드 주의 숲에서 생존했다. 아일랜드의 늑대도 비슷한 과정으로 1770년 마지막 늑대가 죽었다. 스웨덴에서는 순록이 멸종한 이후 늑대가 가축을 먹기 시작하면서 1647년 최초의 늑대 현상금이 도입되었다. 사미인들은 스웨덴 북부에서 조직적으로 늑대를 제거했다. 1960년까지 몇몇 늑대가 스웨덴에서 생존했지만 사냥에 스노모빌을 이용하며 1966년 마지막 늑대가 죽었다. 노르웨이에서는 1973년 마지막 늑대가 죽었다. 핀란드에서는 러시아에서 계속적인 늑대 유입에도 불구하고 20세기 급격하게 줄어들었다. 회색늑대는 1900년 핀란드 동북부 지방에만 서식하다 제2차 세계 대전 시기 개체수가 증가했다.
중앙유럽에서 늑대는 조직적인 사냥과 유제류 개체수 감소로 인해 19세기 초 동안 급격하게 개체수가 감소하였다. 회색늑대는 덴마크에서 1772년 멸종되었고 19세기 전까지 스위스에서 거의 사라졌다. 바이에른주의 마지막 늑대는 1847년 죽었고 라인강 인근에는 1899년 마지막으로 죽었다. 1934년 늑대는 독일 등지에서 이미 멸종한 상태였지만 나치 독일이 늑대 보호 정책을 시작하면서 최초로 늑대를 보호하는 현대적 국가가 나타났다.프랑스의 늑대 사냥은 800년에서 813년 사이 카롤루스 대제에 의해 제도화되었고 "로우베트리에"(louveterie)라는 전문 늑대 사냥 조직을 만든다. 이 조직은 1789년 프랑스 대혁명 이후 폐지되지만 1814년 재조직된다. 1883년에는 1,386마리 이상의 늑대가 죽었다.
동유럽에서는 아시아의 거대 숲 지역에서 유입되는 늑대로 인해 완전히 멸종하지는 않았다. 그러나, 동유럽의 늑대 개체수는 19세기 후반 매우 적은 개체수로 낮아졌다. 슬로바키아의 늑대는 20세기 처음 10년 동안 멸종했고 20세기 중반에는 폴란드 동부의 몇 산림 지역에서만 발견되었다. 동 발칸반도의 늑대들은 넓은 평야, 산, 숲에서 접근성의 혜택을 보았다. 헝가리의 늑대들은 20세기 초반 절반으로 줄었으며 대부분은 판노니아 평원에 몰려 있었다. 루마니아의 늑대 개체수는 1955년부터 1965년동안 매년 4,600마리가 죽으며 1,600마리밖에 남지 않으며 실질적으로 멸종했다. 1967년에는 최소치인 1,550마리까지 줄어들었다. 불가리아에서 늑대의 멸종은 다른 국가보다는 더욱 늦게 이루어져 1955년에는 1,000마리가 남았으며 1964년에는 1-200마리로 줄어들었다. 그리스의 늑대들은 1930년 펠로폰네소스 반도 남부에서 마지막으로 멸종했다. 18세기 동안 강력한 사냥 정책에도 불구하고, 서부 발칸 반도의 알바니아와 유고슬라비아에서는 멸종한 적이 없었다. 늑대의 조직적 사냥은 1923년 유고슬라비아의 슬로베니아 코체베에서 생겨난 늑대 구제위원회(Wolf Extermination Committee, WEC)의 결성으로 시작되었다. WEC는 디나르알프스 산맥에서 늑대 개체수를 줄이는 데 성공했다.
남유럽에서의 늑대는 북유럽과 달리 문화적으로 종의 관용으로 인해 멸종이 완료되지 않았다. 이베리아반도의 늑대 개체수는 1800년대 초 줄어들기 시작했으며 1900년에는 원래의 절반으로 줄어들었다. 이탈리아에서는 늑대에 대한 너그러움으로 인해 1950년 말에 멸종했다. 1800년대 알프스 산맥의 늑대가 사냥당하기 시작하며 1973년에는 100여 마리 정도 남았으며 옛 이탈리아 영토의 3–5% 구역에서 서식하고 있다.
회복

유럽의 늑대 개체수는 전통 전원과 농촌 경제가 축소되며 늑대 박해의 필요성이 줄어들자 1950년 이후 회복이 시작되었다. 1980년대 농촌 지역에서 인구 밀도가 줄어드자 야생 먹이가 증가하며 늑대 개체수가 증가하기 시작했다.
1978년, 스웨덴 중부에서 12년생의 늑대를 재도입하기 시작했고 이후 노르웨이 남부로 확대되었다. 2005년에는 스웨덴과 노르웨이의 늑대 개체수가 11 쌍의 늑대 쌍을 포함하여 백여 마리 이상 서식중인 것으로 파악된다. 회색늑대는 스웨덴에서 완전히, 노르웨이에서 부분적으로 보호된다. 스칸디나비아 늑대의 개체수는 핀란드와 카렐리야 공화국에서 유입되는 늑대들로 인해 증가하고 있다. 핀란드의 늑대는 남부의 3곳에서 보호중이며 특정 계절 및 다른 지역에서만 사냥이 가능하다.
폴란드의 늑대는 1976년 사냥 가능 동물(game species)로 구분되며 이후 8백-9백마리가 개인적으로 서식하고 있다. 폴란드는 중부유럽에서 이웃 국가로 늑대를 확장시키는 주요한 역할을 한다. 북쪽에서는 리투아니아, 에스토니아, 라트비아, 우크라이나와 슬로바키아의 늑대 범위가 겹친다. 2012년 이후에는 폴란드 서부에서 동부로 개체가 이동하며 독일 베를린 15마일(26 km) 이내까지 발견되었다. 슬로바키아의 회색늑대는 보호받지만 예외적으로 가축을 공격할 경우에는 사냥이 허용된다. 몇몇 슬로바키아 늑대가 체코로 이동할 경우에는 완전한 보호를 받는다. 슬로바키아, 우크라이나, 크로아티아의 늑대는 헝가리에서 분산된 것으로 추측되지만, 위치 표지의 부족으로 자율적 개체수 측정이 힘들다. 헝가리의 늑대는 특별한 지위를 부여받고 있지만 문제를 일으킬 경우에는 연중 내내 허가를 받고 사냥을 할 수 있다.
루마니아의 늑대는 약 2,500마리가 있다. 1996년부터 루마니아의 늑대는 법의 적용을 받진 않지만 보호 동물로 지정되었다. 알바니아와 마케도니아 공화국 늑대 개체수는 그리스, 보스니아, 크로아티아의 늑대 경로를 오가는 중요한 곳임에도 불구하고 알 수 없다. 그리스의 늑대는 보호받지만 불법적으로 사냥당하고 있어 전망이 어두운 상태이다. 1986년 이후 보스니아의 늑대는 이웃 국가인 슬로베니아와 크로아티아가 보호 정책을 시작하면서 멸종했다.
이탈리아의 늑대는 1976년 이후 완전히 보호받기 시작했으며, 연간 약 60%가 증가하며 450마리-500마리 정도 서식하고 있다. 이탈리아의 늑대는 1993년 메르칸투르 국립공원 프랑스에서 방사가 시작된 이후로 유입되었으며 2000년 알프스에는 적어도 50마리 이상의 늑대가 서식 중이다. 이베리아반도의 늑대는 2천 마리 정도 있으며 포르투갈 북동부에 150마리가 서식 중이다. 스페인에서는 갈리시아 지방, 레온 주, 아스투리아스 지방에서 서식한다. 이베리아의 늑대는 연간 수백마리가 불법으로 죽고 있지만 개체들은 도루 강 남쪽으로 넘어가 아우스타리우스 동부, 피레네 산맥에도 서식하고 있다.
2011년부터 네덜란드, 벨기에, 덴마크 인접 지역에서 야생에서 이주한 늑대 목격 보고가 이어지고 있다.
아시아
역사적 서식지와 감소
19세기 동안 늑대는 요르단 강 동서의 성지 등에서 서식했다. 그러나, 1964년에서 1980년 동안 농민들의 박해로 개체수가 급감한다. 19세기 동안 사우디아라비아의 북부와 중부로 나누어져 초기에는 대부분 남부인 아시르 주에 서식했고 북부에서는 요르단 산맥 경계와 리야드 주변 지역에 서식했다.
소비에트 연방의 늑대는 거의 모든 지역에 서식하고 있으며 오직 솔로베츠키 제도, 제믈랴프란차이오시파 제도, 세베르나야제믈랴 제도, 카라긴스키섬(Karaginsky Island), 코만도르스키예 제도, 샨타르스키예 제도에서만 서식하지 않는다. 늑대는 적백내전, 제2차 세계 대전 두 시기에 크림 자치 공화국 지역에서 대량 사냥당했다. 두 차례의 세계 대전 이후 소련 늑대 개체수는 두 번 정점을 찍었다. 1940년대 20만 마리에서 3만 마리의 늑대가 죽었지만 연간 4만-5만의 늑대를 낳았다. 소비에트 연방 늑대 개체수는 유럽 러시아에서 많이 사라지며 최저점을 찍었다. 개체수는 1980년 약 75,000마리로 증가했고 1979년에는 32,000마리가 죽었다.내몽골 자치구의 늑대는 1940년 초 먹이인 몽골영양의 밀렵으로 멸종했다.영국령 인도 제국에서는 늑대로 인한 양, 염소, 어린이들의 공격으로 사냥당했다. 1876년에는 노스웨스턴 주와 비하르 주의 늑대 2,825마리가 죽었다. 1920년대에는 노스웨스턴 주와 아와흐(Awadh)에서는 늑대가 절멸했다. 전반적으로 1871년부터 1916년까지 영국령 인도에서 10만 마리 이상의 늑대가 죽었다.
일본의 늑대는 메이지 유신 기간 동안 멸종했고, 이는 "오카미 노 큐조"(ōkami no kujo)로 알려져 있다. 메이지 정부는 늑대가 목장에 위협을 주는 동물로 홍보되었으며 현대 미국의 캠패인과 유사한 늑대 현상금 제도와 화학적 거세를 시행했다. 마지막 일본늑대는 1905년 1월 23일 와시카구시(현재 히가시요시로 알려져 있기도 함)에 사냥당했다.
현대의 서식지

늑대 상태에 대한 신뢰할 수 있는 데이터는 중동의 이스라엘과 사우디아라비아로 안정적인 개체수를 유지하고 있다. 이스라엘은 늑대 보호 조치와 효과적 법 집행으로 적당한 늑대 개체수를 유지하고 있고 이웃 국가로 방출하고 있으며, 사우디아라비아는 3백-6백 마리가 방해받지 않은 광대한 사막 지역에서 서식하고 있다. 사우디아라비아의 늑대는 유목의 감소와 인간 폐기물의 증가로 역사적 범위 내에서 계속 서식하고 있다. 튀르키예의 늑대는 중앙아시아에서 유입되는 개체로 인해 늑대가 유지되고 있다. 튀르키예의 산맥 지역에 산 몇몇 늑대들은 시리아로 옮겨가기도 한다. 작은 늑대 개체가 골란 고원에 서식하고 있으며 군사 활동으로 인해 보호받고 있다. 네게브 사막 남부에 서식하는 늑대는 이집트의 시나이반도나 요르단에서 유입되는 개체로 유지되고 있다. 중동을 통틀어서 늑대를 보호하는 국가는 이스라엘뿐이다. 다른 곳에서는 베두인족이 연중 내내 사냥할 수 있다.
이란에서는 약간의 늑대 개체가 남아 있는 것으로 알려져 있으며, 1970년대 중반 조사로 전국적으로 엹은 밀도로 서식하는 것이 밝혀졌다. 아프가니스탄과 파키스탄 북부는 늑대의 주요 서식지이다. 인도 북부의 잠무 카슈미르 주에는 6만km2에서 3백 마리의 늑대가 서식하는 것으로 알려져 있으며, 히마찰프라데시 주에는 50마리의 늑대가 서식하고 있다. 전반적으로 인도 전역에서 8백-3천 마리의 늑대가 흩어져서 서식하고 있다. 1972년 보호 정책이 시작되지만, 인도늑대는 멸종 위기가 부여되었으며 많은 개체가 점점 감소하고 있으며 인간 거주 지역에 살고 있다. 네팔과 부탄에는 늑대가 서식한다는 어떠한 정보도 없다.
북아시아와 중앙아시아의 늑대 개체는 정확히 알 수 없지만 매년 새끼의 수를 바탕으로 수백-수천 마리로 추산하고 있다. 소비에트 연방의 붕괴 이후 대륙의 늑대 박멸 정책은 중단되었고 전 소련 국가의 늑대 개체수는 25,000-30,000마리로 추산하고 있다. 중국과 몽골에서는 오직 보호구역에서만 늑대가 보호받는다. 몽골의 개체수는 1-3만 마리로 추산되며 중국에서는 더 단편적으로 추산한다. 북부에서는 약 4백 마리의 늑대가 있으며 신장과 티베트에는 1-2천 마리로 추산한다.
한반도의 늑대는 남한의 경우 1980년대를 마지막으로 사라졌다. 그리고 동물원 사육개체도 1997년을 마지막으로 죽어버렸다. 늑대가 사라지면서 현재 그 먹이가 되는 고라니나 노루, 멧돼지와 너구리등 일부 야생동물들이 개체수가 너무 늘어나면서 농작물을 먹어 치워 여러 가지 문제가 발생하였다. 그러자, 일부 동물 보호단체에서 중국에서 만주산 늑대 4마리를 들여와 재도입계획을 세우고 있다. 또한 민간단체에서도 몽골에서 늑대를 들여와 복원을 추진하고 있으며 환경부에서는 북한과 연해주에서 늑대를 도입할 계획이다. 하지만 한반도의 늑대 복원계획에 대해 문제가 있다. 현재 늑대가 복원이 되면 증가하여 여러 문제가 있는 멧돼지나 고라니등 번식력이 좋은 우제류등 야생동물 개체수 조절이 가능하지만 역시 늑대가 사람을 해칠 가능성 역시 제기되고 있다.
북한의 경우에는 많은 지역에서 사라진 것으로 보이며 현재 백두산에 적은 수가 서식한다고 보고되어 있다. 현재 호랑이나 불곰 등 북한의 육식 포유동물이 천연기념물로 지정되어 보호되어 있지만 늑대는 현재 반달가슴곰이나 표범과 마찬가지로 천연기념물에서 배제되어 있다. 아마도 북한에 늑대가 흔하거나 아니면 거의 멸종했을 제기되고 있다.
서울대 황우석 박사는 현재 한국산 늑대를 복제하였다고 논문에 기재하였지만 여러 논란으로 기재를 포기하였다. 그리고 경상북도에서는 멸종위기의 늑대를 복원하기 위해 소백산에 사파리 형식으로 늑대를 방사한다고 한다. 개체들은 몽골이나 러시아에서 들여올 것으로 보고 있다. 또한 대전 동물원의 경우 러시아의 볼가강 유역에서 포획한 늑대들을 도입해 사파리 형식으로 사육한다고 한다. 그 밖에 광주 우치공원이나 청주동물원에서도 한국늑대를 사육하고 있다.
북아메리카
역사적 서식지 및 감소
회색늑대는 원래 북위 20°이북의 모든 지역에 서식지가 있었다. 늑대는 모든 곳으로 퍼져 미국 캘리포니아주 남부와 시에라네바다 산맥 서부, 멕시코 열대 및 아열대 지역으로 확산되었다. 또한, 늑대는 뉴펀들랜드섬, 밴쿠버섬, 알래스카 남동부 섬, 북극 제도, 그린란드 등 주요 섬에서도 서식하고 있다.
북미의 늑대는 인간 인구의 증가와 농업의 확대와 동시에 개체수가 감소하기 시작했다. 20세기 초에 늑대는 애팔래치아 산맥 일부와 오대호 지역 북서부 지역을 제외하고 미국 동부에서 멸종했다. 캐나다에서는 1870년에서 1921년 사이 뉴브런즈윅주와 노바스코샤주에 늑대가 멸종했으며 1911년에는 뉴펀들랜드 주에서 멸종했다. 북부 로키산맥에서 늑대는 1920년대에 사라졌다. 1850년에서 1900년 사이 퀘벡주와 온타리오주 남부에 늑대가 멸종했다. 대평원에서 늑대 개체수 감소는 1860년-70년 사이 아메리카들소 등 다른 먹이의 개체수 감소와 시작되었다. 1900년에서 1930년 사이 종을 멸종하기 위한 집중적 육식 동물 관리 프로그램으로 인해 사실상 미국 서부와 캐나다 국경 인근 부분에서 멸종했다. 회색늑대는 알래스카와 미네소타주 북부를 제외한 모든 연방 및 주에서 멸종되었다. 북미 늑대의 개체수 감소현상은 1930년에서 1950년 사이 남서부 캐나다에서 대형 사냥 게임의 규제와 대형 유제류의 보호로 인해 다시 증가했다. 이 증가는 서부와 북부 캐나다의 늑대 개체수 증가에 도움을 주었다. 1950년대 초부터 1960년대 초까지 늑대 수천 마리가 주로 독에 의해 죽었다. 이 캠패인은 중단되고 1970년대 중반 늑대 개체수는 증가가 시작되었다.

현대의 서식지
그 파장이 미국 서부에도 미치면서 옐로스톤과 아이다호주, 몬태나주에서도 사라지게 되었는데 더그 스미스와 같은 늑대를 연구하는 사람들이 늑대를 재도입하자는 주장이 일었다. 1997년 쯤에 옐로스톤 국립공원과 아이다호 주에 늑대를 재도입을 실행에 옮겼다. 캐나다의 앨버타나 브리티시 컬럼비아에 사는 늑대들을 옐로스톤과 아이다호에서 현지 적응을 마치고나서 방사를 하게 되었다.
옐로스톤 국립공원에서는 늑대가 재도입하기 전까지만 해도 어린 버드나무의 순이 뜯어먹히고 사시나무 또한 그 면적이 줄어들었다. 엘크를 비롯한 초식동물들이 지나치게 불어났기 대문이었다. 하지만 늑대가 재도입되고나서 그러한 문제들이 개선되었으며 사라졌던 비버도 돌아오게 되었다. 그런데 옐로스톤과 아이다호에 재도입된 늑대 무리들간에 세력경쟁이 일어난 적이 있었는데 그 여파로 세력이 약한 여러 무리들이 몬태나 남동부로 가게 되었다. 몬태나 북서부에서는 자연적으로 늑대들이 이주해와서 존재를 알 겨를이 없었다. 옐로스톤의 늑대 재도입은 육식동물 보존 프로그램의 좋은 모델이 되었다.
현대 북미의 늑대 서식지는 미국 알래스카주 및 [[아이다호 주], 몬태나주에도 약간 서식한다. 1995년부터 1996년까지 앨버타주와 브리티시컬럼비아주의 늑대들은 아이다호 주의 옐로스톤 국립공원으로 재도입되었다. 1998년에는 애리조나주와 뉴멕시코주에 재도입되었다. 회색늑대는 캐나다에서 역사적 범위의 80% 이상 서식하고 있어 늑대의 거점으로 꼽히고 있다.
캐나다에서는 캐나다의 행정 구역의 법률에 따라 다르지만 52,000마리에서 6만 마리의 늑대가 서식하고 있다. 캐나다 원주민은 제한없이 늑대 사냥을 할 수 있지만, 다른 주민들은 허가를 필요로 한다. 알래스카 주에서는 6천-7천 마리의 늑대가 서식하는 것으로 추정되며 사냥과 수집철 동안 합법적으로 사냥할 수 있지만 사냥감 제한 등의 기타 제한 법률이 있다. 2002년 기준으로, 옐로스톤의 늑대는 28개 무리 250마리가 있으며 아이다호 주에서는 25개 무리 260마리의 늑대가 서식하고 있다. 1974년부터 위스콘신 주, 미네소타 주, 미시간 주에서는 미국 절멸위기종 보호법에 회색늑대가 포함되었다. 2002년 말에는 에리조나 주와 뉴멕시코 주의 늑대가 8개 무리 28마리까지 줄어들어 멕시코늑대가 절멸위기종 보호법의 보호를 받게 되었다.
2012년 이후 옐로스톤 국립공원에서 옐로스톤 삼각주 무리에 일원이었던 어느 서너살 정도된 수컷이 옐로스톤을 벗어나 사우스다코타주의 파인릿지(소나무산마루)라는 지역으로 이동을 한일이 관측되었다. 전파수신 목걸이로 관측이 되던 개체였는데 이는 로키산맥의 늑대가 동쪽으로 가거나 오대호 연안에 있던 늑대들이 서쪽으로 갈 가능성이 있다는 증거를 제시한 셈이다. 실제로 늑대는 때때로 영토나 짝을 찾기 위해 무리를 벗어나 방랑생활을 하기 때문이다.
아프리카
2011년까지 아프리카에 회색늑대가 서식했는지는 논쟁거리였다. 아리스토텔레스는 이집트에는 그리스에 서식하는 것보다 조금 작은 늑대가 서식한다고 언급했다. 독일의 의학자인 게오르기 엘버스(Georg Ebers)는 현재의 아시우트, 고대 이집트에서는 아누비스라는 도시가 "도시의 늑대"라는 뜻의 이름이 붙어진 것을 근거로 이집트가 늑대를 신성시했다고 주장했다. 동물학자 에른스트 슈와르츠(Ernst Schwarz)는 북아프리카의 황금자칼을 현재의 늑대 같은 개과로 분류했고, 이후 회색늑대와 형태학적 닮음을 간과한 것으로 비판받았다. 2002년 12월, 에리트레아의 다나킬 사막에서 개과 동물이 발견되었으며 황금자칼 또는 주변 지역의 6종과 비슷하지 않고 회색늑대와 비슷한 종을 발견했다.
질병과 기생충
늑대는 먼 거리를 여행하기 때문에 특정 지역에 질병을 전파하고 유지하는 수단이 될 수 있으며 이 중 몇몇은 인간에게 치명적일 수 있다. 그러나, 대부분의 회색늑대에 대해 늑대에 의해 급격하게 발발한 사건은 없다. 보통 질병이 발생한 무리 중 한 늑대는 남겨두고 떠나기 때문에 다른 무리 늑대와의 전염을 방지한다.
바이러스 및 세균 감염
늑대가 감염될 수 있는 바이러스성 질병에는 광견병, 개홍역, 개파보바이러스, 전염성 개간염, 유두종증, 개코로나바이러스등이 있다. 광견병에 감염된 늑대가 주로 분포하고 있는 지역은 러시아, 이란, 아프가니스탄, 이라크, 인도가 있다. 보통 늑대의 잠복기는 8-21일이며 감염 결과 흥분 상태에 이르기 때문에 무리에서 떠나가게 되고, 하루 80 km 이상 움직이기 때문에 다른 늑대에게 전염시킬 수도 있다. 감염된 늑대는 인간에게 두려움을 느끼지 않으며 광견병에 걸린 동물 대부분이 그러듯이 사람을 공격한다. 또한 개에게는 개홍역이 치명적이지만 캐나다와 알래스카 주를 제외하고 늑대를 죽였다는 기록은 나오지 않았다. 개파보바이러스는 대부분 늑대는 살아남지만 새끼 경우에는 탈수로 인한 전해질 불균형이나 내독소로 인한 패혈증으로 사망한다. 늑대는 전염성 개간염에 감염될 수 있지만 실제로 늑대를 사망시켰다는 기록은 없다. 유두종증 경우에는 먹이 동작이 느려질 수 있지만 심각한 질병을 일으키거나 사망하지는 않고 늑대가 사망한 기록은 한 건밖에 없다. 개코로나바이러스는 겨울 몇 달 동안 알래스카 늑대 사이에 심각하게 퍼진 것으로 기록되었다.
늑대가 감염될 수 있는 세균성 질병에는 브루셀라증, 라임병, 렙토스피라증, 툴라레미아, 소결핵균,리스테리아증, 탄저, 구제역 등이 있다. 또한 늑대는 야생 및 사육 순록을 잡아먹으며 돼지브루셀라증(Brucella suis)에 감염될 수 있다. 늑대 성체는 어떤 임상적 징후가 나타나지 않지만, 감염된 암컷은 새끼를 심각하게 약화시킬 수 있다. 라임병은 각각의 늑대를 쇠약하게 만들 수 있지만, 많은 늑대 개체에게 피해를 입히진 않는다. 렙토스피라증은 감염된 먹이나 분비물과의 접촉으로 감염되며 발열, 거식증, 구토, 빈혈, 혈뇨, 황달 등의 증세가 나타나며 심지어는 죽을 수도 있다. 인간 거주 지역 근처에 사는 늑대들은 감염된 가축 폐기물과 부육 등으로 인해 야생의 늑대보다 감염률이 높다. 늑대는 토끼목의 먹이로 인해 툴라레미아에 감염될 수 있지만, 늑대에게 미치는 영향은 알 수 없다. 소결핵균은 늑대에게 주요 위협이 되지 않지만 캐나다에서 두 마리의 새끼를 죽인 기록이 남아 있다.
기생충 감염
늑대는 체외기생균과 채내기생균을 옮기며 이전 소련에서는 최대 50종이 기생균을 옮긴 것으로 기록되었다. 이 기생균의 대부분은 숙주를 아프게 하거나 영양실조에 걸리게 할 수 있지만 대부분은 늑대에게 별다른 증상 없이 옮긴다. 늑대는 개를 통해, 그리고 사람을 통해 기생충을 감염시킬 수 있다. 늑대는 전원 지역에서 살고 기생균을 가축에게 전파시킬 수 있다.
늑대는 벼룩, 진드기, 이목, 응애 등의 다양한 절지동물의 체외기생균에 감염된다. 특히, 대부분의 늑대 새끼는 옴벌레에게 매우 취약하고, 여우와는 달리 심각한 흡윤개선증에 감염된다. 이목 같은 "Trichodectes canis"는 늑대에게 거의 사망의 원인이 될 수 있다. 진드기 같은 익소디즈속은 늑대에게 라임병, 록키산 홍반열 등을 옮긴다.데르마첸토르속(Dermacentor)의 진드기도 늑대를 감염시킨다. 다른 채외기생균에는 이아목을 포함한 이목, 사람벼룩, 개벼룩이 있다.
늑대가 감염되는 채내기생균에는 원생동물과 장내 기생충이 있다(흡충류, 조충류, 선형동물, 구두동물). 3천 종의 원생동물 중 아이소스프라(Isospora), 톡소포자충, 근육포자충, 바베시아, 람블편모충속만 감염시킨다. 늑대는 특히 농부들에게 위험한 네오스포라개구충을 가축들에게 옮기며 감염된 다른 동물들은 유산률이 3-13배 증가한다. 북미 늑대에게서 가장 흔한 알라리아흡혈충은 늑대에게 먹힌 작은 설치류와 양서류에게서 옮긴다. 성숙기에 도달하면 알라리아는 소장으로 이주하고 약간의 증상이 나타날 수 있다. 늑대가 먹은 생선에 의해 감염되는 메토르치스 콘쥰트스는 늑대의 간이나 쓸개에 감염되어 간 질환, 췌장염, 수척해지는 등의 증상이 나타난다. 대부분의 흡충은 늑대의 장에 서식하지만, 웨스트만폐흡충은 폐에 감염된다. 웨스트만흡충은 늑대가 먹은 유제류, 작은 포유류, 물고기를 통해 감염된다. 촌충은 숙주의 면역력과 기생충의 수, 크기에 따라 다르지만 늑대에게 약간의 손상이 가해진다. 늑대가 감염될 시 증상으로는 변비, 독성 알레르기 반응, 점막 자극, 영양실조 등이 있다. 유제류에 의해 감염되는 에키노코쿠스 그라놀로수스는 늑대의 개체 밀도가 증가할수록 감염률이 높아지는 경향이 있는데 늑대는 자신의 대소변으로 에키노코쿠스의 알을 전파시킬 수 있다. 늑대는 30가지 선형동물을 전파시킬 수 있으며 기생충의 수와 숙주의 면역력에 따라 다르지만 대부분 양성 반응을 보인다. 개십이지장충은 늑대의 장에 달라붙어 피를 빨아먹으면서 고색소성빈혈, 수척해짐, 설사 등의 증상이 나타나며 심할 경우 죽기도 한다. 십이지장충으로 알려진 개회충은 자궁을 통해 새끼를 감염시킬 수 있으며 장 자극, 고창증, 구토, 설사 등의 증상이 나타난다. 늑대는 밍크를 통해 거대신충에 감염될 수 있고 신장 질환을 일으키며 이 기생충은 최대 1m까지 자랄 수 있다. 거대신충은 신장 조직의 완전 파괴를 불려오며 양쪽 신장에 감염될 경우 치명적일 수 있다. 늑대는 심장사상충에 감염될 경우 수년간 낮은 수준의 증상이 나타나지만 높은 수준에서는 육두구 간과 심장 팽창 등의 증상을 일으키며 죽을 수 있다. 늑대는 감염된 유제류에 의해 선모충에 감염될 수 있다. 비록 선모충은 늑대에게서 임상 증상을 나타내진 않지만 개에게 수척, 지나친 타액 분비 및 근육 통증을 일으킬 수 있다. 늑대는 구두동물에 거의 감염되지 않지만 러시아에서 오니콜라 스크르쟈비니(Onicola skrjabini), 마크로칸토르휴니쿠스 카투리누스(Macrocantorhynchus catulinus), 모니리포루미스 모니리포루미스(Moniliformis moniliformis) 3종의 감염이 확인되었다.
인간과의 관계

문화
신화와 민속
북유럽 신화와 일본 신화에서 늑대는 신으로 가깝게 묘사되었다. 일본에서는 농부가 신사에서 늑대를 숭배하고 굴 근처에서 먹이를 주며, 야생 맷돼지와 사슴으로부터 작물을 보호하기 위해서 늑대에게 간청한다는 이야기가 있고, 북유럽 신화에서 나오는 펜리르 늑대는 로키의 아들로 묘사되었다. 다른 문화에서, 아일랜드 신화에서는 코르막 맥 에어트가 늑대로 묘사되며,로마 신화에서는 암늑대가 로물루스와 레무스를 기르며 로마를 건국했다는 등 여러 기초 신화에서 늑대를 중심 인물로 두고 있다. 튀르크 신화,, 몽골 신화, 아이누 신화에서는 늑대가 자기 민족의 조상으로 나온 반면,데나이나 민족은 늑대가 한 남성의 형태이며 자신의 형제인 것으로 믿는다. 늑대는 고대 그리스와 고대 로마 신화에서 아폴론 신과 늑대가 태양에 관련되어 있다고 설명하며 북유럽 신화에서는 태양의 신인 스콜이 늑대와 관련되는 등 일부 유라시아의 신화에서 늑대와 태양을 관련시켰다.파웨니 민족의 창조 설화에 따르면 늑대는 죽음을 처음으로 겪은 동물이다. 북유럽 신화에서 볼바 마녀가 히로킨 거인과 힐다 거인을 늑대로 다스리는 것으로 묘사하고, 나바호 족은 늑대를 마녀로 생각하고 두려워하는 등 가끔 북유럽과 아메리카의 일부 원주민 신화에서 늑대는 모두 마법과 관련된 것으로 묘사된다. 마찬가지로, 칠코틴 족은 늑대가 정신 질환과 사망을 일으킬 수 있다고 생각했다.
이솝 우화와 문학

세계에서 가장 오래된 문서 중 하나인 바빌론의 《길가메시 서사시》에서는, 이슈타르 여신이 길가메시를 유혹하자 그녀에게 그녀의 이전 연인들은 양치기가 늑대가 되며 양떼로부터 막지 못하는 동물이 된다는 점을 상기시키고 거부했다.조로아스터교 경전 《아베스타》에서는 늑대가 앙그라 마이뉴 악마의 창조물이며 동물들의 잔인함을 결정한다.아이소포스의 《이솝 우화》에서는 고대 그리스의 정착 시기 늑대가 양 군집 문제를 만드는 것으로 나온다. 이솝 우화 중 가장 유명한 양치기 소년은 의도적으로 거짓말을 하는 것을 지양하는 내용의 동화이며 여기서 관용구 "cry wolf"가 파생되었다. 몇몇의 다른 우화에서는 늑대들은 양치기와 양치기 개, 늑대 간에 대한 경계에 관한 관심을 유지하는 데 집중하며 늑대와 개 사이의 밀접한 관계에 대해 고민을 하는 것으로도 나온다. 아이소포스는 늑대를 이용하여 인간의 도덕적인 행동에 대해 비판했으며, 그의 묘사로 인해 늑대는 위험한 동물이라는 인식이 퍼졌다. 이것은 성경에도 반영되어 늑대가 탐욕스럽고 해악하다는 묘사가 13번 인용되었다.신약성경에서, 예수의 많은 상징으로 이스라엘 문화의 목자 주위에서 늑대가 자기 무리를 보호하듯이 민족을 보호해주는 선한 목자로 묘사했다. 현대의 늑대 이미지는 예수가 양의 가죽을 쓴 늑대를 언급하며 거짓 선지자를 경고하는 것을 비유함으로부터 시작되었다. 몇몇 저자가 늑대를 위험한 사람들과 비교하는 예수의 묘사를 따옴으로서 늑대의 나쁜 인식은 이 때부터 형성되었으며 이후 서구에서 늑대의 탄압에 대한 변명이 되었다. 이후 중세의 기독교 문학에서는 늑대를 성경의 범위에서 더욱 확장시켰다. 7세기 판에 처음 등장하는 《사이올로지스》(Physiologus)에서는 이교도들에게 기독교의 도덕과 신비를 가르친 책이다. 사이올로지스에서는 늑대가 인간을 공격하며 특히 목뼈를 노린다고 적었다. 단테 알리기에리는 암늑대를 사기와 탐욕의 대상으로 나타냈으며 이는 이전에 칸트의 《인페르노》에서 처음 언급되었다. 1487년 처음 출판된 《말레우스 말레피카룸》(Malleus Maleficarum)에서는 늑대는 신이 악인을 처벌하는 대리인이거나 악마의 대리인으로 나와 신도의 믿음을 시험하고 축복을 주는 것으로 언급했다. 12세기 라틴어 시인 예센그리무스(Ysengrimus)에 나오는 아이센그림 늑대는 낮은 귀족을 의미하고, 상대 주인공인 레이너드 여우(Reynard)는 농민의 영웅을 나타냈다. 하지만 충성스럽고 정직하며 도덕적으로 묘사된 아이센그림은 여우의 지혜와 학대의 피해자로 나타나며 종종 이야기 끝에서 죽기도 한다. 1697년 샤를 페로가 처음 쓴 《빨간 두건》(한국에서는 빨간 모자로 알려짐)에서는 서구의 늑대에 대한 부정적인 인식이 다른 문학 작품에 비해 더 많은 영향을 받은 것으로 생각된다. 이 이야기에서는 늑대가 잠재적인 강간범으로 나오며 인간의 목소리를 따라할 수 있다고 적었다.러시아 문학에서는 인간이 눈에 띄게 늑대를 사냥하는 것으로 나오며 이를 따르는 소설을 쓴 작가는 레프 톨스토이, 안톤 체호프, 니콜라이 네크라소프, 이반 부닌, 레오나드 파블로비치 새바네예프 등이 있다. 톨스토이의 전쟁과 평화와 체호프의 "소작농"(Peasants) 두 작품에서는 늑대가 사냥개와 보르조이에게 사냥당하는 장면이 나온다.
팔리 모왓이 1963년 작성한 픽션 회고록 《울지 않는 늑대》(Never Cry Wolf)는 대중 문학에서 처음으로 늑대를 긍정적으로 표현한 작품이며, 또한 늑대에 관한 작품 중 가장 인기있는 작품으로 여겨졌으며 영화화되고 수십년 간 학교에서 가르치기도 하였다. 비록 늑대들을 대중의 인식에서 사랑스럽게 바꿔놓았으나, 사실 부정확하게 협력적이며 귀족적이고 부적절한 지시에 거부하는 등의 늑대에 대한 잘못된 인식을 가져왔다고 비판받기도 하였다. 이 책은 나중에 소비에트 연방 공산당이 늑대의 인간 공격에 관한 올바른 정보 캠패인으로 이용되기도 하였다. 모왓에 관한 이러한 비슷한 비판으로 미사 데포세카의 《미사: 어 메모리제 오브 더 홀로코스트》(Misha: A Mémoire of the Holocaust Years)에서는 폴란드 점령기 동안 늑대가 기른 야생아에 대한 한 저자의 거짓 논란이 나온다.제인 크래이드 조지의 1972년 소설 《줄리에의 늑대》(Julie of the Wolves)의 1부에서는 소녀와 늑대 무리 사이의 관계를 다루고 있다. 이 부의 마지막 항목에서는 늑대의 관점에서 바라보며 의인화되었으나 생물학, 동물학적으로 동물을 바라본다. 여러 현대 아동 문학 작가들은 늑대를 더 긍정적으로 바라보기 위해 고전 동화의 늑대 인식을 고치고 있다. 이 예로 미국의 〈환경적 늑대와 아기돼지 삼형제〉(Eco-wolf and the Three Little Pigs)와 〈양치기 소년〉(The Wolf who cried Boy) 동화가 있다.
문장과 상징

늑대는 영국 병기창 찰지에 자주 이용되었다. 또한, 늑대는 바론 웰비, 바론 렌델, 월셀레이 자작의 장식쇠 서포터와 로베트, 윌슨 대부분의 문장에 볼 수 있다. 데미늑대(demi-wolf)는 많은 울페 성의 가문 구성원의 장식과 문장에 나오는 체르스트에 발에 왕관이 있는 반늑대의 모양으로 나오며, 이는 찰스 2세가 우스터 전투 이후 울페 가문에게 하사한 것이다. 스코틀랜드의 문장, 특히 도나차디쉬 씨족와 스캐니 씨족에 공통적으로 늑대 머리가 나온다. 늑대는 스페인의 문장에서 가장 흔하게 볼 수 있는 것으로 종종 입에 양고기를 물거나 뒤를 건너는 것으로 그려진다. 또한, 늑대는 콜로빌 인디언 연합 동맹과 위스콘신 오나이다 부족, 파위네족의 깃발에서 흔히 볼 수 있다. 현대에서는 늑대가 군사 및 준군사 조직의 문장으로 자주 이용된다. 늑대는 러시아(구 소비에트 연방) 스페츠나츠의 비공식 상징이며 터키 초극단주의 단체인 회색늑대들의 상징이다. 또한 유고슬라비아 전쟁 기간 동안 세르비아의 준군사 조직인 흰 늑대와 부크약늑대에서도 늑대를 상징으로 했다.
가축과 개의 포식

가축 포식은 늑대 사냥의 주요 이유이며 늑대 개체 보존에 심각한 문제를 불러왔다. 이는 늑대의 포식 위협으로 가축 축산에 지장을 주어 경제적 피해를 줄 뿐 아니라 늑대의 박멸로는 이러한 문제를 해결할 수 없다. 일부 국가는 늑대로 인한 피해를 손실 보상 프로그램이나 1963년 처음 제정된 뉴질랜드의 주립 보험 등을 통한 해결책 등을 이용하고 있다.
잘 길들어진 가축은 지속적으로 인간의 보호를 받았기 때문에 자신을 잘 방어할 수 없어서 늑대의 손쉬운 먹이가 된다. 늑대는 일반적으로 야생의 먹이가 떨어질 때 가축 포식으로 의존한다. 유라시아에서는 일부 늑대 개체의 대부분의 식습관이 가축으로 구성되어 있지만, 이러한 문제는 야생의 먹이가 많아진 북미에서는 적은 편이다. 이 가축 손실의 대부분은 늑대 포식에 가장 취약한 여름의 방목 기간 중이나 원격지의 목초지의 방목 가축으로 구성되어 있다. 자주 포식 대상이 되는 가축으로는 양(유럽), 순록(스칸디나비아 북부), 염소(인도), 말(몽골), 소와 칠면조(터키)가 있다. 늑대는 먹이 뒤에서 공격하는 경향이 있기 때문에 말의 강한 뒷발굽 차기 방어수단으로 인해 말보다 늑대가 더 위험할 수 있다. 늑대의 다른 아종들은 우선 다른 동물을 대상으로 포식할 수 있다. 큰 몸집의 북부늑대가 완전히 다 자란 소나 말을 죽일 수 있는 동안 작은 늑대가 큰 늑대에게 맞을 수 있다.
한 번의 공격으로 죽는 동물의 수는 종마다 다르다. 대부분의 가축과 말은 한 번의 공격으로 죽지만, 칠면조, 양 및 순록은 한 번에 죽지 않을 수 있다. 늑대는 주로 동물 방목 시에 공격하기 때문에 때때로 울타리 안으로 넘어가려고 하기도 한다. 큰 몸집의 가축은 늑대에 의해 귀, 꼬리 및 도킹을 포함하여 다리의 밑부분이 다치게 될 수 있다. 몇몇 사례에서, 늑대들은 실제 부정적으로 미치는 경우 가축을 공격하지 않는다. 늑대를 경계하는 가축들은 스트레스로 인한 유산 경험과 체중 감소, 고기 맛 저하 등이 발생한다.
늑대는 경우에는 개를 죽이면서 중요한 음식 공급으로 개에 의존하는 일부 개체가 있다. 일반적으로 늑대는 크기에서 개를 능가하기 때문에 물리적 공격에서 개는 일단 늑대의 큰 머리의 이빨에 물리게 된다. 또한, 늑대와 개가 싸우는 스타일은 다르다. 개는 일반적으로 공격할 시 머리, 목 및 어깨 부근으로 제한되지만 늑대는 상대의 몸과 사지를 문다. 크로아티아에서는 늑대가 양보다 개를 더욱 많이 죽이고 러시아에서는 늑대가 유기견의 개체수를 제한하는 역할을 한다. 개가 사람들과 함께 공격할 때, 늑대는 비정상적인 대담한 행동을 하며 때로는 가까운 사람을 무시하기도 한다. 늑대의 개 공격은 집의 뒷마당이나 숲 모두에서 일어날 수 있다. 마을 외곽에서는 늑대가 개를 기다리기 위해 매복하며 그 뒤에 다른 늑대들로 하여금 개를 유혹하게 하기도 한다. 늑대의 사냥개 공격은 스칸디나비아와 위스콘신 주에서 큰 문제로 대두된다. 스칸디나비아에서 가장 빈번하게 죽은 사냥개는 해리어 품종으로 늙은 동물이 어린 동물보다 소심하여 늑대에 대한 반응을 다르게 할 가능성이 있기 때문에 위험하다. 개의 늑대로 인한 부상은 종종 허벅지와 뒷다리에 나타난다. 치명적인 상처는 보통 목 뒤를 물린 상처이다. 스웨덴 쟘툰드(Jämthund) 같은 대형 개의 사냥은 자신의 방어하기 더욱 수월하기 때문에 살아남을 가능성이 더욱 높다.
인간 공격

늑대의 공포는 인간이 자연적인 늑대의 먹이가 아님에도 불구하고 많은 사회에 만연하고 있다. 늑대가 인간과 마주쳤을 때의 반응은 대부분 이전 경험 유무에 따라 다르다. 인간과의 부정적인 경험이 적거나 식습관이 조절되어 있을 경우 인간에 대한 공포심을 느낀다. 전반적으로 늑대는 적은 수일 경우 보통 인간이 위험하진 않지만, 충분한 먹이와 약간 인간과 접촉했을 경우 가끔 사냥하기도 한다.
늑대는 도발 정도에 따라 반응이 다를 수 있지만 이러한 공격은 주로 즉각적으로 사지를 물면서 공격을 멈출 수 없게 된다. 실제적인 공격은 늑대 한마리가 공격하거나 먹이를 공격하는 방법을 인간을 통해 배우는 것으로 인해 발생한다. 이러한 계속된 공격은 습관화 될 수 있으며 늑대가 인간의 두려움을 잃게 되게 한다. 피해자는 일반적으로 얼굴과 목 주위에 지속적인 공격을 받으며 늑대가 방해가 되지 않는 한 끌려가서 먹힌다. 이러한 공격은 먹이가 죽을 때까지 시간과 공간에 상관없이 계속 공격하는 경향이 있다. 약탈적 공격은 일년 언제나 일어나지만 6-8월 기간에 최고점에 달하며 이때 숲 지역으로 유입되는 사람들(가축 방목이나 식물 채취)로 인해 사람들을 만날 기회가 증가하며 이는 겨울에 광견병에 걸리지 않은 늑대의 공격 사례로 기록되었고 이 공격 지역에는 벨라루스, 러시아 키롭스키 지구, 이르쿠츠크, 카리알라, 우크라이나 지방이 주로 포함된다. 또한, 새끼를 가진 늑대는 이 기간 동안 큰 먹이 스트레스를 받는다. 약탈하는 늑대의 피해자는 거의 18세 미만의 어린이이며 성인의 경우 남성은 매우 드물고 거의 여성이 차지한다.늑대개는 순종 늑대보다 더욱 대담하며 인간의 정착지 주변에 가까이 살기 때문에 따라서 가축과 인간에게 더욱 위협적이다.
광견병의 경우 늑대가 질병의 주요 소유자 역할을 하지 않는 이상 다른 종보다 감염율이 낮지만 개와 여우, 자칼처럼 쉽게 발견되기도 한다. 광견병에 감염된 늑대는 주로 지중해, 중동과 중앙아시아에서 발견되며 북아메리카에서는 매우 드물게 나타난다. 늑대는 매우 분명히 광견병의 "분노" 단계 수준까지 보여줄 수 있다. 이것은 그들의 큰 크기와 힘과 합쳐지기 때문에 광견병에 걸린 늑대가 제일 위험하다. 광견병에 감염된 개보다 늑대가 물렸을 경우 15배 더 높은 것으로 추정된다. 광견병에 감염된 늑대는 보통 혼자 생활하며 먼 거리를 여행하여 종종 많은 가축과 인간을 문다. 대부분 광견병 감염 늑대의 공격은 봄과 가을철에 발생한다. 약탈적 공격과는 달리, 광견병에 걸린 늑대는 먹이를 먹지 않으며 공격은 보통 하루 정도 지속된다. 또한, 대부분의 피해자는 성인 남성이나 보통은 무작위로 선택되어 공격하는 편이다.

늑대의 인간 공격은 드물지만 20세기 이전 유럽에는 가끔 일어나는 일이었다. 프랑스에서는 1580년부터 1830년까지 기간 동안 3,272명이 늑대에 의해 사망했으며 그 중 1,961이 광견병에 감염되지 않은 늑대의 공격이었다. 이탈리아에서는 15세기부터 19세기까지 교회와 정부에 기록된 바로는 440명이 사망했으며, 주로 현대의 스위스 지역인 포 계곡(Po Valley) 지역에 주로 발생했다. 1882년 이전 페노스칸디아 지역은 300년 동안 광견병에 감염되지 않은 늑대에 의해 12세 미만의 아이들 94명이 사망했다.
1840년부터 1861년까지 러시아에서는 광견병에 감염되지 않은 늑대 273마리의 공격으로 성인 7명과 어린이 169명이 사망했다. 1944년부터 1950년까지 러시아 키로프주에서 3-17세의 어린이 22명이 사망했다.(키로프 늑대 공격(Kirov wolf attacks) 참조.) 아시아 대륙의 늑대 공격은 수많은 문서로 보고되었으며 최소 인도에서 수십년 간 거대 늑대 공격이 3번 이상 있었다. 이 공격은 잘 교육된 생물학자가 문서로 기록한 것이 남아 있다.하자리바그흐(Hazaribagh)와 비하르 주에서 1980년부터 1986년까지 늑대의 공격으로 어린이 100명이 사망하고 120명이 부상당했다.
북아메리카 대륙은 매우 낮은 발생률과 기록을 보이고 있으나, 아메리카 토착민들의 구술 역사(oral history)에 따르면 일부 늑대가 인간을 죽였다는 것이 확실시되고 있다. 숲에 사는 부족들은 툰드라에 비해 갑자기 거주지와 가까운 곳에서 늑대가 나타날 수 있으므로 더욱 두려워했다. 이렇게 몇몇의 대형 공격이 유라시아에 비해 북아메리카가 매우 적은 이유는 역사적으로 북아메리카에서 많이 가용된 화기로 인해 유라시아의 늑대보다 북미의 늑대가 인간을 더욱 빨리 두려워했기 했던 것으로 추정된다. 그러나, 북미에서 늑대와의 접촉은 지속적으로 늘어날 것으로 추정한다. 한 연구에서는 알래스카와 캐나다의 늑대 공격은 총 80회로 39회는 매우 건강한 늑대의 습격이었으며 29회는 인간을 전혀 두려워하지 않은 사건으로 판명한다. 북아메리카에서 일어난 광견병에 걸리지 않은 늑대의 대형 습격은 두 차례 있었다. 2005년 캐나다 서스캐처원주에서 한 남성이 사망하였으며, 2010년에는 알래스카 주의 치그니크 호수(Chignik Lake)에서 조깅하던 한 여성이 사망했다.
사냥


늑대는 뛰어난 감각, 빠른 속도와 빠르게 사냥개를 죽일 수 있는 능력, 강한 체력 등으로 인해 늑대 사냥은 매우 어려운 것으로 알려져 있다. 역사적으로 늑대를 사냥하는 많은 방안이 고안되었으며 이는 봄에 둥지에서 태어난 새끼를 죽이는 것을 포함하여 개를 이용한 사냥(보통 시각 하운드, 블러드 하운드, 폭스 테리어를 이용),스트리크닌 중독,과 발판,덫, 러시아에서 인기 있는 사냥 방법인 인간 냄새를 뿌리고 플라드리로 애워싸운 다음 무리 전체를 사냥하는 것이 있다. 이 방법은 늑대의 인간 냄새에 대한 공포에 의존하며 늑대가 냄새에 익숙해지면 효과를 잃을 수 있다. 일부 사냥꾼들은 전화를 이용하여 늑대를 사냥하기도 한다.카자흐스탄과 몽골에서는 전통적으로 늑대를 독수리와 매로 사냥하며 현재는 경험이 풍부한 매잡이가 줄어들어 감소하고 있다. 항공기로 늑대를 쏘는 것은 지상에서보다 가시도가 높기 때문에 매우 효율적이며 이 방법은 논쟁이 있으나 늑대에게 탈출하거나 보호하는 기회를 주기도 한다.
이용

늑대 가죽은 스카프, 여성복의 트림에 사용되며 가끔 재킷, 망토, 외투,머클럭, 융단에도 이용된다. 늑대 가죽은 개가죽보다 얇고 잘 찢어지는 경향이 있다. 늑대가죽의 품질은 모피섬유의 밀도와 강도에 따라 다르고 수직으로 선 털과 측면이 두터운 점이 있다. 이러한 특성은 주로 북부늑대에게 나타나지만 점차 따뜻한 기후로 인해 남쪽으로 옮겨지고 있다. 유라시아늑대보다 북미늑대가 부드럽고 푹신하기 때문에 북미늑대 가죽이 선호되고 가치가 높다. 독에 의해 죽은 늑대의 가죽은 대부분 가치가 높다. 중세 유럽에서는 피부의 악취로 인해 사용되지 않았음에도 불구하고 늑대는 가죽이 유일한 실용적인 측면으로 강조되었다.스칸디나비아 민속에 따르면, 늑대가죽 거들은 입은 사람을 늑대인간으로 변화시키며 여러 아메리카 토착민들은 의약 목적을 위해 늑대가죽을 사용했다.파웨니 족(Pawnee people)은 적 지역을 여행할 때 망토같은 늑대가죽을 입고 다녔다.미국 육군은 제2차 세계 대전과 한국 전쟁에서 병사의 얼굴 동상을 막기 위해 늑대 가죽으로 만든 파카를 입었다. 소비에트 연방에서는 1976년에서 1988년까지 3만 벌의 늑대 가죽이 생산되었다. 멸종위기에 처한 야생동식물종의 국제거래에 관한 협약에 따르면 매년 국제적으로 6천-7천 마리의 늑대가죽이 매년 거래되고 있으며 캐나다, 소비에트 연방, 몽골과 중국에서 가장 많이 수출하며 영국과 미국에서 가장 많이 수입한다. 전반적으로 늑대가죽으로 인한 늑대 개체수 감소는 거의 없지만 북부의 늑대는 개체수 보호를 위해 수입되지 않는다. 늑대 모피는 많은 아메리카 토착민들의 수입으로 이용되고 있다.
늑대가 식품으로 이용되었던 것은 드물지만, 역사적으로 음식이 부족하거나 약용으로 늑대고기를 먹었다. 특히, 1913년 빌하레무르 스테판슨(Vilhjalmur Stefansson)의 북극 원정대는 늑대고기를 많이 섭취했다.자바이칼 원주민들은 먹이가 풍부한데도 불구하고 늑대고기를 섭취했다. 특히 나스카피족(Naskapi)같은 아메리카 토착민들은 늑대고기를 먹을 수 있지만 불충분한 영양이 들어있다고 생각했고 초식동물이 아니었으므로 다른 동식물과 구별되는 특징을 가질 것이라고 생각했다. 아시아의 민간요법에서는 몽골에서는 감기를 완하시키고 늑대고기와 그 폐의 가루를 음식에 뿌려 먹으면 치질이 낫는다고 하는 등 늑대고기와 장기 소비에 주요한 영향을 주었다. 일부 산간 지방의 일본인들은 늑대고기가 용기를 준다고 믿었다. 《더 그레이》 영화 촬영 기간 동안 캐스트 멤버들은 늑대고기를 먹은 것으로 유명했다. 늑대의 고기 맛은 "거칠다"라고 하는 것부터 "연골질 같다", "불쾌하고" "역겹다", "닭고기와 닮았다" "사슴고기와 비슷하다"라는 등 다양한 맛으로 묘사했다.
반려동물로서의 늑대

길들인 늑대는 개보다 예측과 관리가 어렵고 남을 해치는 행위에 대한 이해가 부족하여 완전히 신뢰할 수 없고 약간의 자극에도 사냥 본능(prey drive)을 갖출 수 있다. 그러나 일반적으로 늑대는 다른 개과보다 길들이기 쉬워 새끼부터 오랫 동안 가족과 길들일 수 있고 다른 사람과 보조적인 관계도 가지고 있다. 그러나, 개와는 대조적으로 늑대는 가족을 무리의 연장으로 보고 자신의 생활에서 인정하지만 늑대는 점점 낯선 사람과 무리와 접촉하면 심각한 포비아 현상을 일으킨다. 이에도 불구하고, 미국에서만 늑대를 기르는 인구는 약 8만 명에서 2백만 명에 이르는 것으로 추산되고 혼자 있다는 생각으로 인기가 증가했다. 개는 새끼 때 최대 약 10주까지도 사회화시킬 수 있지만, 늑대는 19일만 지나도 사회화가 거의 불가능하다. 늑대의 모유는 개의 모유보다 더 많은 아르기닌이 들어 있기 때문에 새끼에게 아르가닌을 주사하지 않으면 백내장에 걸릴 수 있다. 개는 쉽게, 그리고 적극적으로 인간 사회와 유대를 유지하지만 늑대는 동종 성체의 부재에서만 유지할 수 있다. 1년 미만의 새끼들은 일반적으로 낯선 사람을 향해 공격하지 않지만 나이가 들며 특히 번식기에는 공격성이 매우 증가한다. 수컷이 암컷보다 공격적이고 다루기 더 힘들다. 늑대는 미국 표준 사육장(kennel)에 넣기 힘들고, 늑대는 개보다 관찰 학습 능력이 뛰어나 단순이 인간이 래치(latch)를 푸는 모습만 보고 따라서 풀 수 있다. 늑대는 한 번 묶여진 상태에서 풀려나는 법을 배우면 다시 묶는 것은 거의 불가능하다.
같이 보기
- 내용주
- 참조주
참고 자료
- (영어) Coppinger, Ray; Coppinger, Lorna (2001). 《Dogs: a Startling New Understanding of Canine Origin, Behavior and Evolution》. Scribner. ISBN 0-684-85530-5.
- (영어) Feldhamer, George A.; Thompson, Bruce Carlyle; Chapman, Joseph A. (2003). 《Wild Mammals of North America: Biology, Management, and Conservation》. JHU Press. ISBN 0801874165.
- (영어) Graves, Will (2007). 《Wolves in Russia: Anxiety throughout the ages》. Detselig Enterprises. ISBN 1-55059-332-3.
- (영어) Harding, Arthur Robert (1909). “Wolf and coyote trapping; an up-to-date wolf hunter's guide, giving the most successful methods of experienced "wolfers" for hunting and trapping these animals, also gives their habits in detail”. Columbus, Ohio, A. R. Harding pub. co.
- (영어) Hemmer, Helmut (1990). 《Domestication: the decline of environmental appreciation》. Cambridge University Press. ISBN 0-521-34178-7.
- (영어) Heptner, V. G.; Naumov, N. P. (1998). 《Mammals of the Soviet Union Vol.II Part 1a, SIRENIA AND CARNIVORA (Sea cows; Wolves and Bears)》. Science Publishers, Inc. USA. ISBN 1-886106-81-9.
- (영어) Lindsay, Steven R. (2000). 《Handbook of Applied Dog Behavior and Training: Adaptation and learning》. Wiley-Blackwell. ISBN 0-8138-0754-9.
- (영어) Linnell, John D. C. (2002). 《The Fear of Wolves: A Review of Wolf Attacks on Humans》 (PDF). NINA. ISBN 82-426-1292-7. 2012년 6월 26일에 원본 문서 (PDF)에서 보존된 문서. 2012년 6월 26일에 확인함.
- (영어) Lopez, Barry H. (1978). 《Of Wolves and Men》. J. M. Dent and Sons Limited. ISBN 0-7432-4936-4.
- (영어) Marvin, Garry (2012). 《Wolf》. Reaktion Books Ldt. ISBN 978-1-86189-879-1.
- (영어) Mech, L. David (1981). 《The Wolf: The Ecology and Behaviour of an Endangered Species》. University of Minnesota Press. ISBN 0-8166-1026-6.
- (영어) Mech, L. David; Boitani, Luigi (2003). 《Wolves: Behaviour, Ecology and Conservation》. University of Chicago Press. ISBN 0-226-51696-2.
- (영어) Pocock, R. I. (1941). “Fauna of British India: Mammals Volume 2”. Taylor and Francis.
- (영어) Roosevelt, Theodore (1909). “Hunting the grisly and other sketches; an account of the big game of the United States and its chase with horse, hound, and rifle” (PDF). New York, London, G. P. Putnam's sons.
- (영어) Seton, Ernest Thompson (1909). 《Life-histories of northern animals : an account of the mammals of Manitoba》 (PDF). New York City : Scribner.
- (영어) Walker, Brett L. (2005). 《The Lost Wolves Of Japan》. University of Washington Press. ISBN 0-295-98492-9.
- (영어) Sillero-Zubiri, Claudio; Hoffman, Michael; MacDonald, David W. (2004). 《Canids: Foxes, Wolves, Jackals and Dogs – 2004 Status Survey and Conservation Action Plan》. IUCN/SSC Canid Specialist Group. ISBN 2-8317-0786-2. 2011년 10월 6일에 원본 문서에서 보존된 문서. 2011년 10월 4일에 확인함.
- (영어) Zimen, Erik (1981). “The Wolf: His Place in the Natural World”. Souvenir Press. ISBN 0-285-62411-3.
- (독일어) Dmitrij Iwanowitsch Bibikow: Der Wolf. Canis lupus (Die Neue Brehm-Bücherei. Bd. 587). 3. Auflage, unveränderter Nachdruck der 2. Auflage 1990. Westarp Wissenschaften, Hohenwarsleben 2003, ISBN 3-89432-380-9.
- (독일어) Patrick Masius, Jana Sprenger: Die Geschichte vom bösen Wolf – Verfolgung, Ausrottung und Wiederkehr. In: Natur und Landschaft. Jg. 87, Nr. 1, 2012, S. 11–16, Abstract Archived 2015년 7월 11일 - 웨이백 머신.
- (독일어) L. David Mech, Luigi Boitani (Hrsg.): Wolves. Behaviour, Ecology, and Conservation. The University of Chicago Press, Chicago IL u. a. 2003, ISBN 0-226-51696-2.
- (독일어) Henryk Okarma: Der Wolf. Ökologie, Verhalten, Schutz. Parey, Berlin 1997, ISBN 3-8263-8431-8.
- (독일어) Ilka Reinhardt, Gesa Kluth: Leben mit Wölfen. Leitfaden für den Umgang mit einer konfliktträchtigen Tierart in Deutschland (BfN-Skripten. 201, ZDB:1476341-2). Bundesamt für Naturschutz (BfN), Bonn 2007, Digitalisat (PDF; 3,3 MB).
- (독일어) Erik Zimen: Der Wolf. Verhalten, Ökologie und Mythos. Das Vermächtsnis des bekannten Wolfsforschers. Neuauflage. Kosmos, Stuttgart 2003, ISBN 3-440-09742-0.
추가 자료
- (이탈리아어) Apollonio, Marco; Mattioli, Luca (2006). 《Il Lupo in Provincia di Arezzo》. Editrice Le Balze. ISBN 88-7539-123-8.
- (러시아어) Bibikov, D. I. (1985). “The Wolf: History, Systematics, Morphology, Ecology”. Nauka, Moscow, USSR. ASIN B001A1TKK4.
-
(스페인어) Blanco, Juan Carlos; Cortés, Yolanda (2001). 《Ecología, censos, percepción y evolución del lobo en España: análisis de un conflicto》. Sociedad Española para la Conservación y Estudio de los Mamíferos (SECEM). ISBN 84-607-3896-5.
|title=에 외부 링크가 있음 (도움말) -
(영어) Boitani, Luigi (2000). 《Action Plan for the conservation of the wolves (Canis lupus) in Europe》. Council of Europe Publishing.
|title=에 외부 링크가 있음 (도움말) - (영어) Busch, Robert H. (2009). 《Wolf Almanac》. The Lyons Press. ISBN 1-59921-069-X.
- (영어) Coleman, Jon T. (2006). 《Vicious: Wolves and Men in America》. Yale University Press. ISBN 0300119720.
- (영어) Dutcher, Jim; Dutcher, Jamie (2003). 《Wolves at Our Door: The Extraordinary Story of the Couple Who Lived with Wolves》. William Andrew. ISBN 0-7434-0049-6.
- (영어) Fischer, Hank (1995). 《Wolf Wars》. Falcon. ISBN 1560443529.
- (영어) Fuller, Todd K. (2004). “Wolves of the World”. Voyageur Press. ISBN 0896586405.
- (영어) Grooms, Steve (1999). “Return of the Wolf”. Northword Press. ISBN 1559717173.
- (영어) Hampton, Bruce (1997). 《The Great American Wolf》. Holt Paperbacks. ISBN 0805055282.
- (영어) Harrington, Fred H.; Paquet, Paul C. (1982). 《Wolves of the world: perspectives of behavior, ecology, and conservation》. Simon & Schuster. ISBN 0-8155-0905-7.
- (영어) Mech, L. David (1993). “The Way of the Wolf”. Voyageur Press. ISBN 0896581799.
- (영어) McIntyre, Rick (1996). 《A Society of Wolves: National Parks and the Battle over the Wolf》. Voyageur Press. ISBN 0896583252.
- (영어) McNamee, Thomas (1998). 《The Return of the Wolf to Yellowstone》. Holt Paperbacks. ISBN 0805057927.
- (영어) Musiani, Marco; Boitani, Luigi; Paquet, Paul C. (2010). 《The World of Wolves: New Perspectives on Ecology, Behaviour, and Management》. University of Calgary Press. ISBN 1-55238-269-9.
- (영어) Nie, Martin (2003). 《Beyond Wolves: The Politics of Wolf Recovery and Management》. University of Minnesota Press. ISBN 0300119720.
- (러시아어) Pavlov, Mikhail P. (1982). “The Wolf in Game Management”. Agropromizdat, Moscow.
- (독일어) Kontaktbüro Wolfsregion Lausitz, Internationaler Tierschutz-Fonds (Hrsg.): Wölfe vor unserer Haustür. Im Grenzgebiet zwischen Deutschland und Polen. 2014, (PDF; 1,3 MB).
- (독일어) Kontaktbüro Wolfsregion Lausitz, Internationaler Tierschutz-Fonds (Hrsg.): Wenn Sie einem Wolf begegnen. 2014, (PDF; 1,0 MB).
- (독일어) Staatsministerium für Umwelt und Landwirtschaft des Freistaats Sachsen (Hrsg.): Förderung des präventiven Herdenschutzes im Wolfgebiet. 3. Auflage. Dresden 2011, (PDF; 353 kB).
- (독일어) Stefan Willeke: Die Wölfe kommen, in: Die Zeit, 1. April 2015, S. 11–13
외부 링크
- (영어) California Wolf Center(캘리포니아 늑대 센터)
- (영어) 당신이 늑대를 접할 경우(If You Encounter a Wolf), Viltskade Center (2003년) 제작.
- (영어) The International Wolf Center(국제 늑대 센터)
- (영어) IUCN 늑대 종 그룹 안내
- (영어) 늑대 서식 지역에서의 안전 숙박 방법, Alaska Department of Fish and Game (2009년 1월)
- (영어) UK Wolf Conservation Trust(영국 늑대 보호 신탁)
- (영어) 전설의 죽음(Death of a Legend) 와 야생의 울음(Cry of the Wild), 빌 마슨(Bill Mason) 제작.

| 종 및 아종 |
|
 |
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 역사 | ||||||||||||
| 생태 | ||||||||||||
| 문화 | ||||||||||||