
외재성 막 단백질(外在性膜蛋白質, 영어: peripheral membrane protein)은 관련된 생물학적 막에 일시적으로만 부착되는 막 단백질이다. 이들 단백질은 통합막 단백질에 부착되거나 지질 이중층의 주변 영역을 관통한다.
예를 들어, 많은 이온 채널과 막관통 수용체의 조절 단백질 소단위는 외재성 막 단백질로 정의될 수 있다. 통합 막 단백질과 달리 외재성 막 단백질은 단백질 정제과정에서 추출된 모든 단백질의 수용성 성분 또는 분획에 모이는 경향이 있다. GPI 앵커가 있는 단백질은 이 규칙의 예외이며 통합 막 단백질과 유사한 정제 특성을 가질 수 있다.
생물학적 막에 대한 단백질의 가역적 부착은 다양한 메커니즘을 통해 세포 신호 및 기타 많은 중요한 세포 이벤트를 조절하는 것으로 나타났다. 예를 들어, 많은 효소와 생물학적 막 사이의 밀접한 연관은 그것들을 지질 기질과 가깝게 만들 수 있다.막 결합(membrane binding)은 또한 많은 단백질 구조 도메인 내에서 재배열, 해리 또는 구조적 변화를 촉진하여 생물학적 활성을 활성화할 수 있다. 또한, 많은 단백질의 위치는 상주하는 막의 내부 또는 외부 표면 또는 소엽에 국한된다. 이것은 적절한 단백질-단백질 상호작용의 가능성을 증가시켜 다중 단백질 복합체의 조립을 용이하게 한다.

지질 이중층과의 결합

외재성 막 단백질은 다른 단백질과 상호작용하거나 지질 이중층과 직접 상호작용할 수 있다. 후자의 경우, 그들은 양친매성 단백질로 알려져 있다.G-단백질 및 특정 단백질 키나아제와 같은 일부 단백질은 막횡단 단백질 및 지질 이중층과 동시에 상호작용한다. 일부 폴리펩티드 호르몬, 항균 펩티드 및 신경독은 세포 표면 수용체 표적을 찾아 상호작용하기 전에 막 표면에 축적되며, 이는 그 자체가 외재성 막 단백질일 수 있다.
세포 표면 막을 형성하는 인지질 이중층은 친수성의 두 영역 사이에 끼워진 소수성 내부 코어 영역으로 구성된다. 하나는 내부 표면에, 다른 하나는 외부 표면에 있다. 세포막). 모델 인지질 이중층의 내부 및 외부 표면 또는 계면 영역은 약 8 ~ 10Å의 두께를 갖는 것으로 나타났지만, 이는 다량의 강글리오사이드 또는 지질다당류를 포함하는 생물학적 막에서 더 넓을 수 있다. 전형적인 생물학적 막의 소수성 내부 코어 영역은 SAXS(Small angle X-ray scattering)에 의해 추정되는 약 27 내지 32 Å의 두께를 가질 수 있다. 소수성 내부 코어와 친수성 계면 영역 사이의 경계 영역은 약 3Å로 매우 좁다(성분 화학 그룹에 대한 설명은 지질 이중층 문서 참조). 소수성 코어 영역에서 바깥쪽으로 이동하고 계면 친수성 영역으로 이동하면 물의 유효 농도가 이 경계층을 가로질러 거의 0에서 약 2M의 농도로 빠르게 변한다. 인지질 이중층 내의 인산염 그룹은 물로 완전히 수화되거나 포화되어 소수성 코어 영역의 경계 외부에서 약 5 Å에 위치한다.
일부 수용성 단백질은 지질 이중층과 비가역적으로 결합하여 막횡단 알파-나선 또는 베타-배럴 채널을 형성할 수 있다. 이러한 변형은 colicin A, alpha-hemolysin 등과 같은 기공 형성 독소에서 발생한다. 그들은 또한 BcL-2 유사 단백질, 일부 양친매성 항균 펩타이드 및 특정 아넥신에서 발생할 수 있다. 이러한 단백질은 구조적 상태 중 하나가 수용성이거나 막과 느슨하게 결합되어 있기 때문에 일반적으로 외재성로 설명된다.
막 결합 메커니즘
단백질과 지질 이중층의 결합은 단백질의 3차 구조 내에서 상당한 변화를 수반할 수 있다. 여기에는 이전에 펼쳐진 단백질 구조 영역의 접힘 또는 접힘의 재배열 또는 단백질의 막 관련 부분의 재 접힘이 포함될 수 있다. 또한 단백질 4차 구조 또는 올리고머 복합체의 형성 또는 해리, 이온, 리간드, 조절 지질의 특이적 결합이 포함될 수 있다.
전형적인 양친매성 단백질은 생물학적 기능을 수행하기 위해 지질 이중층과 강하게 상호작용해야 한다. 여기에는 지질 및 기타 소수성 물질의 효소 처리, 막 고정, 서로 다른 세포막 사이의 작은 비극성 화합물의 결합 및 전달이 포함된다. 이러한 단백질은 이중층과 단백질 표면의 노출된 비극성 잔기 사이의 소수성 상호작용, 조절 지질과의 특정 비공유 결합 상호작용 또는 공유 결합 지질 앵커에 대한 부착을 통해 이중층에 고정될 수 있다.
많은 외재성 단백질의 막 결합 친화도는 이들이 결합된 막의 특정 지질 조성에 의존하는 것으로 나타났다.
비특이적 소수성 결합
양친매성 단백질(amphitronic protein)은 다양한 소수성 앵커 구조를 통해 지질 이중층과 연결된다. 양친매성 α-나선, 노출된 비극성 루프, 번역 후 아실화 또는 지질화된 아미노산 잔기, 또는 포스파티딜이노시톨 포스페이트와 같은 특이적으로 결합된 조절 지질의 아실 사슬과 같은 . 소수성 상호 작용은 천연 소수성 앵커가 존재하는 경우 MARCKS 단백질 또는 히스타토필린의 다염기성 도메인과 같은 높은 양이온성 펩타이드 및 단백질에 대해서도 중요한 것으로 나타났다.
지질 고정 단백질
지질 고정 단백질(Covalently bound lipid anchors)은 팔미 토일화, 미리스토일화 또는 프레닐화를 통해 세포막의 세포질 측면에 있는 서로 다른 지방산 아실 사슬에 공유적으로 부착된다. 세포막의 소포체 표면에서 지질 고정 단백질은 지질 글리코실포스파티딜이노시톨(GPI) 및 콜레스테롤에 공유적으로 부착된다. 아실화 잔기의 사용을 통한 막과의 단백질 결합은 가역과정이다. 아실 사슬은 막에서 해리된 후 단백질의 소수성 결합 주머니에 묻힐 수 있기 때문이다. 이 과정은 G 단백질의 베타 소단위 내에서 발생한다. 아마도 구조적 유연성에 대한 이러한 추가 필요성 때문에 지질 앵커는 일반적으로 단백질 결정학 연구에 의해 잘 해결되지 않는 단백질 3차 구조의 매우 유연한 세그먼트에 결합된다.
특정 단백질-지질 결합

일부 세포질 단백질은 주어진 막 내에서 발견되는 특정 유형의 지질을 인식하여 다른 세포막으로 모집된다. 특정 지질에 대한 단백질의 결합은 단백질 내에서 발생하고 이들이 결합하는 지질의 지질 헤드 그룹에 대한 특정 결합 포켓을 갖는 특정 막 표적화 구조 도메인을 통해 발생한다. 이것은 전형적인 생화학적 단백질- 리간드 상호작용이며 분자간 수소결합, 반데르발스 상호작용 및 단백질과 지질 리간드 사이의 소수성 상호 작용의 형성에 의해 안정화된다. 이러한 복합체는 또한 칼슘 이온(Ca 2+)을 통해 단백질의 아스파르테이트 또는 글루타메이트 잔기와 지질 인산염 사이의 이온성 다리 형성에 의해 안정화된다. 이러한 이온성 다리는 이온(Ca 2+와 같은)이 지질 결합 이전에 용액에서 단백질에 이미 결합되어 있을 때 발생할 수 있고 안정적이다. 이온성 다리의 형성은 단백질 C2 유형 도메인과 아넥신 사이의 단백질-지질 상호작용에서 볼 수 있다. .
단백질-지질 정전기적 상호작용
양전하를 띤 단백질은 비특이적 정전기적 상호작용에 의해 음전하를 띤 막에 끌리게 된다. 그러나 모든 외재성 펩타이드와 단백질이 양이온은 아니며 막의 특정 면만 음전하를 띠고 있다. 여기에는 원형질막의 세포질 측, 외부 세균막 및 미토콘드리아 막의 외부 전단지가 포함된다. 따라서 정전기적 상호작용은 사이토크롬 c와 같은 전자 운반체의 막 표적화, 카리브도 톡신과 같은 양이온성 독소 및 일부 PH 도메인, C1 도메인 및 C2 도메인과 같은 특정 막 표적화 도메인에서 중요한 역할을 한다.
정전기적 상호 작용(Electrostatic interaction)은 용액의 이온 강도에 크게 의존한다. 이러한 상호 작용은 생리학적 이온 강도(0.14M NaCl)에서 상대적으로 약한다. 시토크롬 c, 카리브도 톡신 또는 히사토 필린과 같은 작은 양이온성 단백질의 경우 ~3~4kcal/mol이다.
막의 공간적 위치
막에 있는 많은 양친매성 단백질 및 펩티드의 방향 및 침투 깊이는 부위 지향 스핀 라벨링, 화학적 라벨링, 단백질 돌연변이의 막 결합 친화도 측정,형광 분광법, 용액 또는 고체 상태를 사용하여 연구된다. NMR 분광법, ATR FTIR 분광법, X선 또는 중성자 회절, 및 계산 방법.
단백질의 두 가지 별개의 막 결합 모드가 확인되었다. 일반적인 수용성 단백질에는 노출된 비극성 잔류물이나 기타 소수성 앵커가 없다. 따라서, 그들은 완전히 수용액에 남아있고 에너지적으로 비용이 많이 드는 지질 이중층으로 침투하지 않다. 이러한 단백질은 이중층과 정전기적으로만 상호 작용한다. 예를 들어 리보뉴클레아제 및 폴리-리신은 이 모드에서 막과 상호 작용한다. 그러나, 전형적인 양친매성 단백질은 계면 영역을 관통하여 막의 탄화수소 내부에 도달하는 다양한 소수성 앵커를 갖는다. 이러한 단백질은 지질 이중층을 "변형"하여 지질 유체-겔 전이 온도를 감소시킨다. 결합은 일반적으로 강한 발열 반응이다. 멤브레인과 양친매성 α-나선의 결합도 유사하게 발생한다. 비극성 잔기 또는 지질 앵커가 있는 본질적으로 구조화되지 않거나 전개 되지 않은 펩티드는 특히 이러한 펩티드가 양이온성이고 음으로 하전된 막과 상호작용할 때 막의 계면 영역을 관통하여 탄화수소 코어에 도달할 수 있다.
분류
효소
외재성 효소는 지질(포스포리파제 및 콜레스테롤 산화효소), 세포벽 올리고당(글리코실 트랜스퍼라제 및 트랜스글리코시다제) 또는 단백질(신호 펩티다제 및 팔미토일 단백질 티오에스테라제)과 같은 다양한 막 구성요소의 대사에 참여한다. 리파아제는 또한 물에서 미셀 또는 비극성 방울을 형성하는 지질을 소화할 수 있다.
| 수업 | 기능 | 생리학 | 구조 |
|---|---|---|---|
| 알파/베타 가수분해효소 접힘 | 화학 결합의 가수분해를 촉매한다. | 박테리아, 곰팡이, 위 및 췌장 리파아제, 팔미토일 단백질 티오에스테라아제, 큐티나아제 및 콜린에스테라아제 포함 | |
| 포스포리파제 A2 (분비 및 세포질) | 인지질의 sn-2 지방산 결합의 가수분해 . | 지질 소화, 막 파괴 및 지질 신호 전달 . | |
| 포스포리파제 C | 포스파티딜 이노시톨인 PIP2를 2개의 두 번째 메시지인 이노시톨 트리포스페이트 및 디아실글리세롤 로 가수분해한다. | 지질 신호 | |
| 콜레스테롤 산화효소 | 콜레스테롤을 cholest -4-en-3-one으로 산화 및 이성질화한다. | 박테리아 병인에 사용되는 콜레스테롤의 세포막을 고갈시킨다. | |
| 카로티노이드 옥시게나제 | 카로티노이드를 절단한다. | 카로티노이드는 식물과 동물 모두에서 호르몬 (인간의 비타민 A 포함), 색소, 향료, 꽃 향기 및 방어 화합물로 기능한다. | |
| 리폭시게나제 | 고도불포화 지방산의 이산 소화를 촉매하는 철 함유 효소. | 동물에서 리폭시게나제는 류코트리엔으로 알려진 염증 매개체의 합성에 관여한다. | |
| 알파 독소 | Phospholipase C와 유사하게 세포막에서 인지질을 절단한다. | 박테리아 병인, 특히 Clostridium perfringens . | |
| 스핑고미엘리나제 C | 포스포디에스테라아제는 포스포디에스테르 결합을 절단한다. | 스핑고미엘린과 같은 지질 처리. | |
| 글리코실 트랜스퍼라제: MurG 및 트랜스글리코시다제 | 활성화된 공여체 분자에서 특정 수용체 분자로 당 부분의 이동을 촉매하여 글리코시드 결합을 형성한다. | 이당류, 올리고당 류 및 다당류 (당접합체)의 생합성 MurG는 세균성 펩티도글리칸 생합성에 관여한다. | |
| 페로킬라타제 | 프로토포르피린 IX를 헴으로 변환한다. | 포르피린 대사에 관여하는 프로토포르피린은 달걀 껍질을 강화하는 데 사용된다. | |
| 근튜불린 관련 단백질군 | PtdIns3P 및 PtdIns(3,5)P2를 탈인산화하는 지질 포스파타제 . | 근육 세포 분화에 필요하다. | |
| 디하이드로오로테이트 탈수소효소 | 디하이드로오로테이트(DHO)를 오로테이트로 산화 . | 원핵 및 진핵 세포에서 피리미딘 뉴클레오티드의 생합성. | |
| 글리콜산 산화효소 | α- 하이드록시산이 상응하는 α- 케토산으로 산화 되는 것을 촉매한다. | 녹색 식물에서 효소는 광호흡에 참여한다. 동물에서 효소는 옥살산 염 생산에 참여한다. |
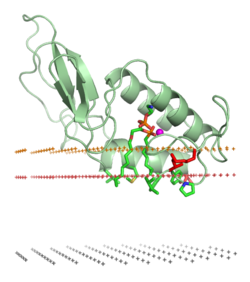
막 표적화 도메인("지질 클램프")
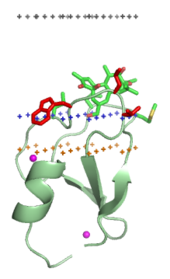
막 표적화 도메인(membrane-targeting domain)은 막에 내장된 지질 리간드의 헤드 그룹과 특이적으로 연관된다. 이러한 지질 리간드는 서로 다른 유형의 생물학적 막에 다양한 농도로 존재한다. 예를 들어, PtdIns3P는 초기 엔도솜의 막에서 주로 발견되고, 후기 엔도솜에서는 PtdIns(3,5)P2, 골지체에서는PtdIns4P이다. 따라서 각 도메인은 특정 멤브레인을 대상으로 한다.
- C1 도메인[1]은 디아실글리세롤과 포르볼에스테르에 결합한다.
- C2 도메인[2]은 포스파티딜세린, 포스파티딜콜린 또는 PtdIns(3,4)P2 또는 PtdIns(4,5)P2에 결합한다.
- Pleckstrin 상동성 도메인[3], PX 도메인[4] 및 Tubby 도메인[5]은 서로 다른 포스포이노시티드에 결합한다.
- FYVE 도메인[6]은 PtdIns3P에 대해 더 구체적이다.
- ETH 도메인[7]은 PtdIns(3,4)P2 또는 PtdIns(4,5)P2에 결합한다.
- ANTH 도메인[8]은 PtdIns(4,5)P2에 결합한다.
- ERM(ezrin/radixin/moesin) 계열의 단백질[9]은 PtdIns(4,5)P2에 결합한다.
- 다른 포스포이노시티드 결합 단백질에는 포스포티로신 결합 도메인[10]과 특정 PDZ 도메인이 포함된다. PtdIns(4,5)P2를 바인딩한다.
- 혈액 응고 인자의 디스코이딘 도메인[11]
- ENTH, VHS 및 ANTH 도메인[12]
구조 도메인
구조 도메인(structural domain)은 막에 대한 다른 단백질의 부착을 매개한다. 멤브레인에 대한 결합은 annexin 또는 GLA 도메인에서와 같이 산성 단백질 잔기와 지질의 인산염 그룹 사이에 다리를 형성하는 칼슘 이온(Ca2+)에 의해 매개될 수 있다.
| 수업 | 기능 | 생리학 | 구조 |
|---|---|---|---|
| 별관 | 칼슘 의존성 세포내막/ 인지질 결합. | 기능에는 소포 트래피킹, 막 융합 및 이온 채널 형성이 포함된다. | |
| 시냅신 I | 시냅스 소포를 덮고 여러 세포골격 요소에 결합한다. | 신경 전달 물질 방출 조절 기능. | |
| 시누클레인 | 알 수 없는 세포 기능. | 원형질막의 안정성 및/또는 회전율을 조절하는 역할을 하는 것으로 생각된다. 파킨슨병과 알츠하이머병 모두와 관련이 있다. | |
| 응고 시스템의 GLA 도메인 | 감마-카르복시글루타메이트 (GLA) 도메인은 칼슘 이온의 고친화도 결합을 담당한다. | 혈액 응고 캐스케이드에서 응고 인자의 기능에 관여한다. | |
| 스펙트린 및 α-액 티닌 -2 | 여러 세포골격 및 미세섬유 단백질에서 발견된다. | 원형질막 완전성 및 세포골격 구조의 유지. |
작은 소수성 분자의 운반체
이러한 외재성 단백질은 서로 다른 유형의 세포막 사이 또는 막과 세포질 단백질 복합체 사이에서 비극성 화합물의 운반체 역할을 한다. 수송된 물질은 포스파티딜이노시톨, 토코페롤, 강글리오사이드, 당지질, 스테롤 유도체, 레티놀, 지방산, 물, 거대분자, 적혈구, 인지질 및 뉴클레오티드이다. 이하의 내용은 영문 위키를 그대로 기술한다.
- Glycolipid transfer proteins
- Lipocalins including retinol binding proteins and fatty acid-binding proteins
- Polyisoprenoid-binding protein, such as YceI protein domain
- Ganglioside GM2 activator proteins
- CRAL-TRIO domain (α-Tocopherol and phosphatidylinositol sec14p transfer proteins)
- Sterol carrier proteins
- Phosphatidylinositol transfer proteins and STAR domains
- Oxysterol-binding protein
전자 캐리어
이 단백질은 전자 수송 사슬에 관여한다. 여기에는 시토크롬 c, 쿠프레독신, 높은 잠재성 철 단백질, 아드레노독신 환원효소, 일부 플라보단백질 등이 포함된다.
폴리펩티드 호르몬, 독소 및 항균 펩티드
많은 호르몬, 독소, 억제제 또는 항균성 펩티드가 막횡단 단백질 복합체와 특이적으로 상호작용한다. 그들은 또한 단백질 표적에 결합하기 전에 지질 이중층 표면에 축적될 수 있다. 이러한 폴리펩티드 리간드는 종종 양전하를 띠고 음이온성 막과 정전기적으로 상호작용한다.
일부 수용성 단백질과 펩티드는 또한 막횡단 채널을 형성할 수 있다. 그들은 일반적으로 올리고머화(oligomerization), 상당한 구조적 변화(conformational changes)를 겪으며, 막과 비가역적으로 결합한다. 그러한 막횡단 채널 중 하나인 α-헤모리신의 3D 구조가 결정되었다. 다른 경우에, 실험 구조는 주변에서 지질 이중층과 상호작용하는 수용성 형태를 나타내지만, 채널 형성 펩티드의 일부는 다소 소수성이므로 유기 용매 또는 미셀의 존재하에서 NMR 분광법에 의해 연구되었다.
| 수업 | 단백질 | 생리학 |
|---|---|---|
| 독 독소 |
|
잘 알려진 종류의 생물독소에는 신경독, 세포 독, 헤모톡신 및 괴사 독소가 있다. 생물독소는 포식(뱀, 전갈의 독소)과 방어(꿀벌 및 개미 독소)의 두 가지 주요 기능이 있다. |
| 말미잘 독소 | 나트륨 및 칼륨 채널의 억제와 막 기공 형성은 알려진 40종 이상의 말미잘 펩타이드 독소의 주요 작용이다. 말미잘은 육식 동물이며 포식과 방어에 독소를 사용한다. 말미잘 독소는 가장 독성이 강한 유기인산염 화학무기와 유사한 독성을 가지고 있다. | |
| 박테리아 독소 | 미생물 독소는 다양한 병원성 박테리아의 주요 독성 인자이다. 일부 독소는 세포막을 용해시키는 기공 형성 독소이다. 다른 독소는 단백질 합성을 억제하거나 2차 메신저 경로를 활성화하여 다양한 세포 기능을 유지하는 데 중요한 신호 전달 경로에 극적인 변화를 일으킨다. 여러 박테리아 독소는 초항원으로 작용하고 면역 체계를과도하게 확장시키는 대규모 T 세포 증식을 일으켜 면역 체계에 직접 작용할 수 있다. 보툴리눔 독소는 신경 분비 소포가 신경 시냅스 원형질막과 도킹/융합하는 것을 방지하여 신경 전달 물질 방출을 억제하는 신경 독소이다. | |
| 곰팡이 독소 |
|
이들 펩타이드는 특이한 아미노산인 α-아미노이소부티르산의 존재를 특징으로 하며, 멤브레인 채널 형성 활성으로 인해 항생 및 항진균 특성을 나타낸다. |
| 항균 펩타이드 |
|
항균 펩타이드가 박테리아를 죽이는 작용 방식은 다양하며 세포막 파괴, 대사 방해, 세포질 성분 표적화를 포함한다. 많은 기존의 항생제와 대조적으로 이 펩타이드는 정균성(bacteriostatic) 대신 살균성인 것으로 보이다. |
| 방어기제 |
|
Defensin은 일종의 항균성 펩타이드이다. 미생물 침입에 대한 거의 모든 타고난 숙주 방어의 중요한 구성 요소이다. Defensin은 전기적 인력을 통해 미생물 세포막을 관통하고 막에 기공을 형성하여 유출을 허용하여 궁극적으로 미생물을 용해시킨다. |
| 신경 펩티드 |
|
이 단백질은 뉴런을 자극하고 행동 반응을 유발하며 강력한 혈관 확장제이며 다양한 유형의 평활근 수축을 담당한다. |
| 아폽토시스 조절제 |
|
Bcl-2 계열의 구성원은 미토콘드리아 외막 투과성을 관장한다. Bcl-2 자체는 림프구 및 신경 세포를 포함한 다양한 세포 유형에서 세포자멸사를 억제한다. |