

미토콘드리아(영어: mitochondria 단수:mitochondrion[*]), 고대 그리스어 mitos: 끈 + chondros: 낱알)는 진핵생물에서 산소 호흡의 과정이 진행되는 세포 속에 있는 중요한 세포소기관으로, 한자 표기로는 사립체(絲粒體) 또는 활력체(活力體)라고도 한다. 미토콘드리아라는 이름은 아래의 모식도에서 잘 나타나 있듯이 겉모양이 낱알을 닮고 내부 구조가 마치 끈을 말아 놓은 것 같다고 하여 붙여진 이름이다.
기본적인 기능이 여러 유기물질에 저장된 에너지를 산화적 인산화 과정을 통하여 생명활동에 필요한 아데노신삼인산(ATP)의 형태로 변환하는 것이기 때문에 미토콘드리아는 세포의 발전소라고 할 수 있다. 보통 미토콘드리아는 세포의 25%의 세포질을 차지하고 있으나 그 크기와 수가 세포의 종류와 역할에 따라 다양하다. 자체적인 DNA(미토콘드리아 DNA)와 RNA를 가지고 있어 세포질 유전에 관여한다. 자체적인 DNA의 존재와 이중막 구조는 미토콘드리아뿐만 아니라 엽록체에서도 나타나는 것으로 오래전 세균에 의한 세포내 공생의 결과로 진핵생물의 탄생이 이루어진 데서 유래한 것으로 여겨진다.
개요
미토콘드리아에는 여러 가지 모양의 것이 있다는 사실이 전자 현미경의 연구로 알려져 있으나, 광학 현미경에서는 아주 작은 과립으로밖에 보이지 않는다. 하나의 세포 내에 존재하는 미토콘드리아의 수는 세포에 따라 다르지만, 보통 많은 수가 존재한다. 전자 현미경에 의한 관찰에 따르면, 미토콘드리아는 바깥쪽을 둘러싸는 막질 부분과 내부로 이루어져 있다. 막 부분은 외막과 내막의 이중막으로 되어 있는데, 내막의 일부는 안쪽으로 돌출하여 여러 겹으로 접혀 있다. 이렇게 접혀 있는 내막의 구조를 ‘크리스테(cristae)’ 또는 크리스타(crista)라고 한다. 미토콘드리아에는 TCA회로나 전자 전달계에 관여하는 모든 효소가 들어 있다. 따라서, 미토콘드리아는 세포내 호흡에 의해 유기물의 화학에너지를 ATP로 바꾸는 중요한 기관이다.
구조

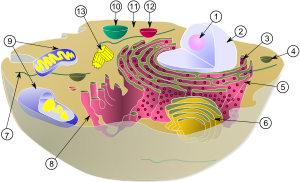
1. 외막
2. 막 사이 공간
3. Intercristal space
4. Peripheral space
5. 라멜라 구조
6. 기질
7. 크리스테 구조
8. 내막
9. Inner boundary membrane
10. Cristal membrane
11. Mitochondrial DNA
12. Matrix granule
13. 리보솜
14. ATP 합성 효소
15. Porin
미토콘드리아는 세포막과 같은 형태가 2중으로 겹친 2중막 구조로 이것은 단백질을 포함하고 있는 인지질 2중층으로 이루어진 내막과 외막으로 이루어져 있다. 외막과 내막의 기능은 각각 다르다. 내막 내부를 기질(matrix)이라고 부르며 내막과 외막의 사이를 막간공간(intermembrane space)이라고 한다. 한편 미토콘드리아의 길이는 약 2~6 마이크로미터 정도이며, 직경은 약0.5마이크로미터(μm) 정도이다.
외막
미토콘드리아 전체를 감싸고 있으며 무게 기준으로 인지질과 단백질이 반반씩 구성하고 있다. 포린(porin)(2~3 nm 정도 크기의 비교적 큰 내재성단백질(integral proteins))이 많이 있는데 이 포린은 5000D(달톤)이하인 분자가 투과할 수 있는 통로(channel)이다. 5000D(5KD)보다 큰 분자들은 능동수송에 의해서만 외막을 통과할 수 있다. 지방산의 탄화수소부분의 합성/신장, 에피네프린(아드레날린)의 산화, 트립토판의 분해등의 기능에 관련된 다양한 효소들이 이러한 능동수송 과정에 작용한다.
내막
4가지 종류의 기능을 가진 단백질(효소)을 가지고 있다.
- 산화효소: 세포호흡
- ATP 생성효소:아데노신삼인산/ATP생성
- 내막 안팎으로의 대사물질운송
- 단백질 수송
내막은 구불구불 접혀져 있기 때문에 막 표면적이 넓어져 ATP생성 능력을 높여준다. 이러한 내막이 늘어난 부분을 크리스타(crista)라고 하고, 이런 구조형태를 '크리스타 구조'라고 한다. 예를 들어 간의 미토콘드리아는 크리스타를 포함한 내막 표면적이 외막의 5배에 달하며 근육세포와 같이 더 많은 ATP를 요구하는 세포의 경우 간세포의 경우보다 더 많은 크리스타를 가지고 있다
미토콘드리아 기질
기질(matrix)은 내막으로 둘러싸인 공간으로 미토콘드리아, 리보솜, tRNA, DNA, 게놈의 복사본과 수백 가지의 효소가 존재하고 있다. 효소들의 기능은 지방산과 피루브산(pyruvate)의 산화, TCA회로를 포함한다. 미토콘드리아는 자체적인 유전물질을 가지고 있으며 자체적으로 RNA와 단백질을 만들 수 있다. 핵에서 유래하지 않은 미토콘드리아 DNA는 내막을 구성하는 펩티드(사람에서는 13개의 펩티드)정보를 저장하고 있다. 자체 생성된 펩티드는 숙주세포의 핵에서 유래한 폴리펩티드와 함께 미토콘드리아 막을 구성한다.
미토콘드리아의 기능
미토콘드리아의 기본적인 기능은 유기물질을 세포가 사용하는 에너지 형태인 ATP로 전환하는 것이다.
미토콘드리아의 일부 기능은 특정한 세포에서만 수행된다. 예를 들어 간세포의 미토콘드리아는 단백질 대사의 부산물인 암모니아를 해독하는 효소를 가지고 있다. 이러한 기능을 조절하는 유전자의 변이는 여러 가지 미토콘드리아 질병을 일으킬 수 있다.
에너지 전환
미토콘드리아의 주 기능은 ATP의 생산으로 해당과정(미토콘드리아의 바깥쪽 세포질에서 이루어진다)의 주생산물인 피루브산(Pyruvic acid)과 NADH 대사를 통하여 이루어지거나 또는 지방산의 베타산화(β oxidation) 과정에서 얻어진다. ATP의 생성은 세포의 유형이나 산소의 존재유무에 따라 2 가지 방법이 있다.
TCA회로(tricarboxylic acid cycle)
해당과정에서 생성된 피루브산 분자는 능동수송에 의해 미토콘드리아 내막을 거쳐 기질로 들어간다. 기질에 들어간 피루브산은 coenzyme A에 결합해 acetyl CoA를 생성하고 생성된 acetyl CoA는 TCA회로(구연산회로 또는 Krebs 회로)로 들어가게 된다. 1개의 피루브산에 의해 3개의 NADH와 1개의 FADH2가 생성되어 전자전달계에 관여하게 된다. 미토콘드리아 내막에 붙어있는 숙신산 탈수소효소(succinate dehydrogenase)를 제외하고 TCA회로에 관여하는 모든 효소는 미토콘드리아 기질에 녹아 있다. 한편 지방산은 베타산화(beta酸化)를 통해 지방산이 미토콘드리아에서 연속적으로 산화하는 현상으로 이러한 단계를 거쳐 에너지를 획득할 수 있다.
화학삼투와 산화적 인산화
TCA회로에서 반응후 생성된 NADH와 FADH2가 미토콘드리아 기질에서 미토콘드리아 내막에 있는 전자전달계 단백질(ComplexⅠ~Ⅳ)중 NADH는 complexⅠ 에게 고에너지 전자 2개를 주면서 NAD+로 산화된다. 그렇게 전자를 얻은 ComplexⅠ에서 전자 친화력(전자를 끌어당기는 힘)이 높은 구조로 인해 고에너지 전자가 위에서부터 아래로 계단을 타고 내려오듯 전달된다. 전달될 때 전자의 에너지가 손실되며 그 손실된 에너지를 이용해 전자전달계의 일부 복합체들은 수소이온(H+)을 기질에서 막간공간으로 2개씩 수송한다. ComplexⅠ에서 전달이 끝난 전자는 Coenzyme-Q가 전자를 받아 ComplexⅢ로 전달한다, 동시에 FADH2는 ComplexⅡ에 전자를 전달하여 FAD로 산화된다. 또한 ComplexⅠ 과 비슷한 양상으로 전자를 전달시키고, ComplexⅠ과 같이 Coenzyme-Q가 ComplexⅢ로 전자를 전달한다. 이때 ComplexⅡ는 직접적으로 수소이온을 통과시키지 않으며, complexⅢ와 complexⅣ의 수소이온 통과에 있어 관여한다. 전자를 가지고 ComplexⅢ로 간 Coenzyme-Q는 전자를 하나는 통과시켜 Cytochrome-C로 전달하고, 다른 하나는 다시 Coenzyme-Q에 되돌아간다. ComplexⅢ에서 Cytochrome-C로 전달된 전자는 ComplexⅣ로 전달된다. 전자를 받은 ComplexⅣ는 Cytochrome C 2개로부터 전달받은 2개의 전자를 전자의 최종 수용체인 O2 와 두개의 수소이온이 반응해 물(H2O)가 만들어진다. 결과적으로 막간공간과 미토콘드리아 기질의 수소이온의 농도 기울기가 생겨 ATP합성효소(ATPase)가 막간공간에 있는 수소이온을 미토콘드리아 기질로 통과시키면서 120º가량 회전하면서 ADP와 Pi(무기인산)을 ATP로 합성시킨다

복제와 유전
미토콘드리아는 세포의 에너지 요구에 반응하여 DNA를 복제하고 분열한다. 세포의 에너지요구량이 높아지면 미토콘드리아는 신장하여 분열하며 에너지 요구량이 낮아지면 미토콘드리아는 파괴되거나 불활성화 상태가 된다. 세포가 분열할 경우 세포질의 분배가 다소 불균등하기 때문에 미토콘드리아의 분배 또한 균등하지 않다. 미토콘드리아는 세균과 같이 이분법으로 자신을 복제하지만 세균과는 달리 미토콘드리아끼리 융합하기도 한다. 때로는 단백질과 폴리리보솜이 많은 곳에서 새로운 미토콘드리아가 생기기도 한다.
미토콘드리아의 유전자는 핵의 유전자와 같은 방법으로 유전되지 않는데, 정자에 의한 난자의 수정에서 난자의 핵과 정자의 핵이 동등하게 수정란의 세포핵유전자에 기여하는 반면 미토콘드리아는 난자의 것만 유전된다. 난자에 수정되는 정자는 단 1개뿐이며 정자의 미토콘드리아는 편모를 움직이는 에너지를 내는 데 사용되다 난자에 들어오게 되면 이내 파괴되게 된다. 반면 난자 자체는 정자에 비해 상대적으로 적은 미토콘드리아를 가지고 있지만 살아남아서 분열을 계속하여 성체의 세포에 존재하게 된다. 즉 미토콘드리아는 거의 대부분 모계(母系)유전하는 것이다.
미토콘드리아 DNA의 모계유전(maternal inheritance)은 동물을 포함 대부분의 진핵생물에서 볼 수 있다. 드물게 부계유전을 하는 경우도 있는데 침엽수(소나무와 주목 제외)에서 흔하게 나타나며, 인간의 경우 아주 낮은 확률로 나타난다고 한다.
단일계통 유전은 서로 다른 미토콘드리아 계통 간의 유전자 재조합 가능성이 낮고 미토콘드리아 DNA가 대체로 클론임을 의미한다. 만약 유전자 재조합이 없다면 모든 미토콘드리아 DNA는 일배체형(haplotype, 홑가닥)으로서 동일하게 유전되기 때문에 집단의 진화적 역사를 연구하는 데 유용하게 사용될 수 있다. 하지만 종종 인간의 미토콘드리아 DNA 상호간에는 재조합이 일어나기도 한다.
미토콘드리아는 미토콘드리아가 유래되었을 것으로 추정되는 진정세균 무리보다 무척 적은 규모의 게놈을 가지고 있다. 일부는 소실된 것으로 보이지만 상당수가 세포핵으로 이전한 것으로 보인다. 이러한 현상은 진화적으로 볼 때 비교적 일반적으로 보인다.
미토콘드리아의 단일계통 유전은 일부 효모종의 미토콘드리아의 Petite 돌연변이(자신의 DNA 일부 또는 모두를 잃어 산화적 인산화를 못하는 비정상적인 미토콘드리아 돌연변이. petite와 정상적인 효모사이의 융합이 일어나게 되면 딸세포의 일정 비율은 petite가 됨.)에서 볼 수 있듯이 유전자 간 분쟁(intragenomic conflict)을 야기할 것으로 보인다. 이러한 세포소기관의 분쟁은 남성과 여성의 분화로 해결할 수 있으며 유전자간 분쟁은 성의 분화가 일어난 원인 중 하나로 생각된다.
집단 유전학에서의 이용
미토콘드리아 DNA는 유전자 재조합이 거의 없기 때문에 집단 유전학이나 진화 생물학을 연구하는 과학자들에게 유용한 자료를 제공해준다. 반수체의 형태, 즉 거의 대부분 모계로부터 유전되기 때문에 다른 개체간의 미토콘드리아 DNA의 관계는 계통수(系統樹, phylogenetic tree)로 나타낼 수 있으며 미토콘드리아 DNA의 유형분석을 통해 집단의 진화역사를 추론해 볼 수 있다. 인간 진화 유전학에서 대표적인 예가 아프리카기원설(인류 최초의 여성조상 미토콘드리아 이브의 추적)과 네안데르탈인 연구이다.
앞서 언급했듯이 미토콘드리아는 거의 모계로만 유전되어 재조합이 잘 일어나지 않기 때문에 최초의 여성인류를 추적할 수 있다. 모든 인류에게서 미토콘드리아 DNA 유형이 거의 일치하기 때문에 인류의 아프리카 기원설을 뒷받침하는 근거가 되고 있다.
네안데르탈인의 경우 해부학적으로 현생인류와 공통점이 없어 현생인류의 조상인지 여부에 대하여 논란이 있었지만 미토콘드리아 DNA 연구 결과 현생인류와 연관성이 없음이 밝혀져 네안데르탈인 후예의 존재여부에 대한 논란에 종지부를 찍게끔 하였다. 그러나 미토콘드리아 DNA는 집단에서의 여자(모계)쪽의 역사만을 반영하기 때문에 전체 군집의 역사를 보여주지 못한다는 단점을 안고 있다. 예를 들어 인구의 분산이 주로 남자에 의해 이루어졌다면 미토콘드리아 연구로는 알아내지 못한다. Y염색체(포유동물에서 재조합이 없는 영역)를 포함한 핵DNA연구(재조합으로 분석하기가 힘듦)로 보다 넓은 범주의 집단의 진화적 역사를 연구한다.
미토콘드리아는 리보솜과 GOZA를 가지고 있고 스스로 분열할 수 있기 때문에 독립적인 생활을 하던 원핵생물의 세포 내 공생으로 유래했다고 본다. 하지만 구체적인 기원에 대해서 의견이 분분하다. 미토콘드리아 DNA는 대체로 원형이고 여러 유전정보를 담고 있어 원시세균 무리 중 특수하게 분화된 것이 원시 미토콘드리아로 되었다는 가설도 있고(Futuyma 2005), 리케차(rickettsia)와 연관되어 있을 것이라는 가설이 있지만 세포내 공생에서 유래했다 점에서는 동일하다. 세포 내 공생에 관해서도 기생에 의한 것이라는 이견도 있지만 숙주의 내포작용에 의한 것이라는 의견이 지배적이다. 원시 미토콘드리아의 숙주세포 내 공생으로 숙주는 세포호흡을 할 수 있게 되어 같은 영양을 공급받더라도 효율적으로 사용할 수 있게 되었고 원시 미토콘드리아는 생존을 보장받고 양질의 양분(당)을 공급받게 되어 상호간의 상승작용이 일어나 생존할 수 있는 영역의 확장이 일어났을 것으로 추측된다. 미토콘드리아뿐만 아니라 엽록체 또한 세포내 공생으로 상승작용이 일어난 것으로 보고 있다.
적어도 20억 년 전 세포공생이 이루어졌다고 추측되는 미토콘드리아는 아직도 그 기원에 대한 증거를 가지고 있다. 첫째, 미토콘드리아는 진핵세포(80S 리보솜)와 달리 원핵생물과 유사한 70S(침강계수, sediment coefficient)리보솜을 가지고 있다. 둘째, DNA가 원핵생물과 유사하다. 반복구간이 없이 코딩DNA비율이 높고 대체로 인트론(intron)이 없으며 원래 원핵생물보다 적은 유전정보를 담고 있지만 직선이 아닌 원형DNA를 가지고 있다는 점이 유사하다. 미토콘드리아는 인트론이 거의 없고 반복구간이 없음에도 불구하고 원형 mRNA를 잘라 다시 접합(polyadenylation)하여 완성된 mRNA를 만드는 다인자 전사(multigenic transcript)를 한다.
일부 진균(microsporidian)이나 원생생물(metamonad, entamoebid, 독립영양 pelobiont)과 같은 진핵생물에서는 미토콘드리아가 없는 경우도 있다. rRNA계통수에서 이들은 매우 원시적인 진핵생물로 여겨지며 미토콘드리아의 세포공생 이전에 출현한 것으로 보는 시각도 있으나 미토콘드리아에 의해 유도된 세포소기관 과 유전정보를 가지고 있으므로 실제로 미토콘드리아가 없는 진핵생물은 없다고 봐야한다.
결론적으로 미토콘드리아의 존재는 진핵생물의 발달에서 중요한 시발점이라 할 수 있겠다.
당뇨병
미토콘드리아의 에너지 대사기능에 비추어볼때 당뇨병(DM)등의 메커니즘 이해에서 운동이 세포내 미토콘드리아의 발생 기작과 매우 주요한 관계가 있다고 보고되는 일련의 과정이 알려져있다는 점은 매우 주요한 정보를 시사한다.
같이 보기
- 물질대사(신진대사)
- 화학삼투설
- 엽록체
- 전기화학 퍼텐셜
- 세포 내 공생설
- 해당과정
- 미토콘드리아 질병
- 미토콘드리아 DNA
- 미토콘드리아 유전학
- 미토콘드리아 투과성 변이공(Mitochondrial permeability transition pore, Mt PT pore)
- 생활습관병
- 대사 증후군(메터볼릭 신드롬)
- 만성 피로 증후군(CFS)
- 인슐린 민감성(IS) VS 인슐린 저항성(IR)
참고 자료
- Futuyma, Douglas J. (2005). “On Darwin's Shoulders”. 《Natural History》 114 (9): 64–68.
- Scheffler, I.E. (2001). “A century of mitochondrial research: achievements and perspectives”. 《Mitochondrion》 1 (1): 3–31.
- Alberts, Bruce; et.al. (1994). 《Molecular Biology of the Cell》 Thi판. New York: Garland Publishing Inc.