

텔로미어(telomere) 또는 말단소립 (末端小粒)은 염색체의 끝부분에 있는 염색 소립으로 세포의 수명을 결정짓는 역할을 한다. 이것은 즉 세포시계의 역할을 담당하는 DNA의 조각들이다. 텔로미어는 그리스어의 '끝'(τἐλος, telos)과 '부위'(μέρος, meros)의 합성어다. 세포분열이 일어나는 동안에 염색체와 DNA를 복제하는 효소는 염색체의 끝부분으로 복제를 계속할 수 없다. 텔로미어가 없는 상태로 세포가 분열된다면 세포에 관한 정보가 들어있는 염색체의 끝부분이 소실될 것이다. 텔로미어는 염색체의 끝부분을 막고있는 분해되지 않는 완충지역이라 할 수 있다. 하지만 세포가 분열되면서 텔로미어는 소실되며 텔로머레이스라는 역전사효소에 의해 보충된다.
발견
1970년대 초, 러시아 이론가 알렉세이 올로브니코프(Alexei Olovnikov)가 그 자신의 끝 부분을 완전하게 복제해내지 못하는 염색체들을 발견했다. 이 발견과 헤이플릭(Hayflick)의 개념에 기초하여 DNA의 일부는 세포의 수명이 끝날 때까지 DNA 복제 시 계속 소실될 것이라 가설을 세웠다. 그러나, 그의 예상은 세포 노화와 불멸화에 대해 연구하는 일부 연구자에게만 알려졌다.
1975년에서 1977년 사이에 엘리자베스 블랙번(Elizabeth Blackburn)는 단순히 염색체 말단을 구성하는 DNA 조각을 가지고 비범한 텔로미어의 특성을 발견했다. Blackburn과 Carol Greider, 그리고 Jack Szostak은 염색체가 어떻게 텔로미어와 텔로머레이스 효소에 의해 보호되는지 밝혀 노벨상(Physiology of Medicine)을 수상했다.
그럼에도 불구하고, 1970년대에는 텔로미어가 줄어드는 메커니즘이 보통 세포 분열 횟수에 제한을 둔다는 인식은 전혀 없었을 뿐만 아니라, 이러한 현상이 세포 노화에 큰 영향을 줄 것이라고 제시하는 연구도 없었다. 또한, 그 메커니즘이 수명을 제한 시킨다는 인식도 없었다.
이후, 생명공학 기업인 Geron이 세포 노화에서 텔로미어 축소에 어떻게 영향을 미치는지 밝히기 위해 RNA와 텔로머레이스에 대하여, 세포 불멸화(cell immortalization) 상태에서 텔로머레이스의 재활성화에 대해 지속적으로 투자를 했다.
텔로미어의 연구 과정
세포의 노화에 대해서 구체적으로 연구한 Leonard Hayflick 박사는 1961년, 생물과 장기에 따라서 세포의 분열 횟수가 정해져 있고, 그 후에 세포가 노화해 죽는다는 사실을 밝혀냈다. Hayflick 박사는 태아의 세포는 100번 정도 분열하고, 노인의 세포는 20~30번 정도 분열한 후에 노화가 된다는 사실을 발견했다. 이를 헤이플릭 리미트(Hayflick Limit)라고 불린다. Hayflick 박사의 연구에 의하면 고양이는 8번, 말은 20번, 인간은 60번 정도 세포분열을 할 수 있다고 한다. 그 후에 발견된 것이 바로 텔로미어이다. 1990년대 초가 되어서야 생물세포학자들에 의해서 텔로미어가 염색체의 말단에 위치함이 밝혀졌다. 연구가 계속 진행된 결과, 샌프란시스코 캘리포니아 대학(UCSF)의 Elizabeth blackburn(61)교수를 비롯하여 존스홉킨스 의대 Carol Greider(48)과 하버드 의대 Jack Szostak(57)은 텔로미어를 통해서 세포의 노화 메커니즘을 규명하였다. 이들은 2009년 노벨 생리학·의학상 수상자로 선정되었다.
구조
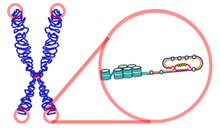
말단
텔로미어의 말단은 3`말단이 단일가닥으로 되어있는데, 이 부분은 t-loop(telomeric-loop)를 구성함으로써 안정된 구조를 형성한다. 이 안으로 묻히게 되는 구조를 'end-capping' 구조라 일컫는다. 이중가닥 중 튀어나온 한 가닥을 G-strand라 하는데, 이는 치환이 되어(displace) 'D-loop(displacement-loop)'를 형성하고 있다. 이러한 구조들은 염색체 말단 구조를 보호하기 위한 것으로 여겨진다. 그러나 텔로미어가 융합되면서 두 개의 동원체를 갖는 염색체(dicentric chromosome)를 만들 수 있고, 이는 궁극적 유전자 재배열을 일으켜 유전체의 불안전성을 초래할 수 있다.
텔로미어-연합단백질(telomere-associated proteins)
텔로미어 결합 단백질(telomere-binding proteins)

현재까지 포유류에서 발견된 텔로미어 DNA에 염기서열 특이적으로, 직접적으로 결합할 수 있는 텔로미어 단백질은 TRF1, TRF2, POT1이다.
| 텔로미어 DNA 서열에 결합하는 단백질 | |
|---|---|
| TRF1 | 텔로미어 길이 유지의 음성적 조절(negatively regulated) |
| TRF2 | 텔로미어 말단 보호에 중요한 조절 역할 |
| POT1 | 단일가닥 DNA 부분인 텔로미어 TTAGGG overhang에 결합 |
쉘터린(shelterin) 복합체
텔로미어에 직접적으로 결합하지는 못하지만(TRF1, TRF2, POT1과 달리) 텔로미어 단백질 복합체에 참여할 수 있는 단백질도 있는데 이를 텔로미어 연합 단백질이라 한다. 위 언급한 세 단백질 포함하여 RAP1, TPP1, TIN2 등이 그 예다. 이들을 쉘터린 복합체라 일컫는다. 텔로미어에 쉘터린 복합체가 결합해 온전한 텔로미어를 구성하면 텔로미어에서 DNA 수선 기작이 작동하여 재조합을 일으키는 것을 막아준다. 또한, 텔로머레이스가 텔로미어로 접근하려 하는 것을 저지한다. TIN2는 텔로미어 DNA 서열에 직접적으로 결합하지는 못하지만, 그 대신 다른 단백질인 TPP1과의 결합으로 TRF1 및 TRF2와 POT1을 이어주는 다리 역할을 한다. TPP1은 POT1과 POT1-TPP1 복합체를 형성한다(TPP1은 텔로미어 DNA 서열에 직접적으로 결합하지 못한다).
| 텔로미어 연합 단백질(인간) | 기능(추정) | 텔로미어 결합 자리 | |
|---|---|---|---|
| 이중가닥 | 단일가닥 | ||
| TRF1 | 텔로미어 길이 조절, 기능 유지 | o | |
| TRF2 | 텔로미어 말단 보호, 텔로미어 길이 조절 | o | |
| Ku70/80, DNA-PK 복합체 | DNA 손상 수선, 텔로미어 길이와 구조 조절 | o | |
| Tankyrase (TANK1, TANK2) | TRF1의 ADP 리보실화를 통한 텔로미어 길이 조절(positively regulate telomere length) | DNA에 직접 결합
못하고 TRF1과 상 호 작용함으로써 참여 |
|
| TIN2(TRF-interacting protein 2) | TRF1의 구조적 기능을 중재(negatively regulate telomere length) | TRF1과 상호 작용 | |
| RAP1(repression and activation protein 1) | 텔로미어 길이 조절(positively regulate telomere length) | TRF2와 상호 작용 | |
| POT1 | 텔로미어 길이 조절, 텔로미어 말단 보호, 단일가닥 overhang(돌출부분)부분의 보호 | o | |
| hTERT | 텔로머레이스의 촉매 소단위(catalytic subunit) | ||
| Ku | DNA 수선(repair) | TRF1, TRF2와
상호작용 |
|
| MRN(MRE11/RAD50/NBS1) | 세포주기확인점(cell cycle checkpoint) 조절, DNA 수선 | TRF2와 상호 작용 | |
| PinX1 | 텔로머레이스 억제 | TRF1, 텔로머레이
스와 상호 작용 |
|
TRF1이나 TIN2를 과발현 시킬 경우 텔로미어 길이가 짧아진다.
열린 구조와 닫힌 구조
닫힌 구조 상태의 텔로미어는 텔로머레이스가 텔로미어 길이를 신장 시키는 것을 방해하고, 열린 구조 상태의 텔로미어는 텔로머레이스와의 결합을 쉽게 하여 텔로미어 길이 증가를 유도할 수 있다. 쉽게 말해 텔로미어가 닫힌 상태여야 안정된 상태라는 것이다. 쉘터린 구조는 텔로머레이스의 텔로미어에 대한 접근 가능성을 간접적으로 제한시켜 텔로미어 길이의 음성적(negative) 조절에 관여한다. 따라서 쉘터린 단백질들은 '닫힌 텔로미어 구조' 또는 'cap의 형성(capping)'을 촉진하고, 텔로미어 단일가닥 돌출부분(3`-overhang)에 텔로머레이스가 접근하는 것을 방지한다.
성별에 따른 텔로미어 길이의 차이
여성의 텔로미어가 남성이 가지는 것보다 더 길다고 밝혀졌다. 서던 법(Southern blot)을 통해 이와 같은 결론을 도출해 냈으나, 이 발견은 아직 방법론적 차이가 있는 설명이 더 필요하다.
텔로미어의 작용
텔로미어의 길이는 종에 따라서 매우 다양하다. 효모에서는 300~600개의 염기쌍으로 이루어져있고, 인간의 경우 수 킬로베이스(DNA 등 핵산 연쇄의 길이 단위)로 이루어져 있다. 인간의 경우 텔로미어의 길이가 짧아짐에 따라서 세포분열을 막는 노화현상을 유발할 수 있다. 텔로미어는 6개(인간의 경우)의 특이적인 DNA 염기서열이 수백에서 수천 번 반복되며, 염색체의 말단에 위치하고 있어서 세포가 분열할 때 염색체가 분해되는 것을 막아준다. 텔로미어의 특징을 살펴보면, 세포가 한 번 분열할 때마다 염색체 말단으로부터 50~200개의 텔로미어 DNA 뉴클레오타이드를 잃어버린다. 텔로미어의 길이가 짧아질수록 세포가 늙었다는 것을 의미한다. 그렇기 때문에 여러 차례 세포분열을 하면서 대부분의 텔로미어 DNA가 손실되면 세포는 세포분열을 멈춘다.
텔로미어 염기서열
| 생물체 | 염기서열 (5'에서 3'을 향함 ) |
|---|---|
| 척추동물(vertebrate)
사람 생쥐(Mice) 쥐(Rats) 조류 |
TTAGGG |
| 붉은빵곰팡이 | TTAGGG |
| 애기장대 | TTTAGGG |
| 클라미도모나스 | TTTTAGGG |
| 누에나방 | TTAGG |
| 효모 | TTAC(A)(C)G(1-8) |
텔로머레이스

손실되는 텔로미어의 DNA를 복구하는 효소가 존재하는데 그것을 텔로머레이스(말단소립 복제효소, telomerase)라고 한다. 이 효소 덕분에 세포가 분열해도 텔로미어의 길이를 어느 정도의 길이로 유지할 수 있다. 효소가 지나치게 활성화되면 세포가 계속 분열할 수 있다. 진핵 세포로 이루어진 생물체에서 텔로머레이스가 활발한 세포는 소장 내부의 표피세포(상피세포), 골수세포, 암세포 등이다. 소장 내부의 표피세포는 끊임없이 음식물, 체액과 접촉하면서 상처를 입거나 떨어져 나가곤 한다. 하지만, 활성화된 텔로머레이스에 의해서 세포분열이 지속적으로 일어나면서 상처입거나 떨어져 나간 표피세포를 보충할 수 있는 것이다.
텔로머레이스는 텔로미어의 DNA 염기서열과 상보적인 염기쌍을 가지는 RNA를 이용하여 텔로미어 DNA를 만들어내는 일종의 역전사 효소를 만들어 낼 수 있다. (RNA의 상보적인 염기쌍 : C(Cytosine)-G(Guanine), A(Adenine)-U(Uracil)) 예를 들어서 역전사효소의 RNA의 염기서열이 AAUCCC이면 TTAGGG를 갖는 텔로미어 DNA염기서열이 만들어지는 것이다. 이와 같은 방법으로 텔로머레이스는 염색체 말단의 텔로미어의 길이가 짧아지지 않도록 해준다.
텔로미어의 축소(세포 노화)

DNA 복제가 DNA 말단에서부터 시작되지 않고 중앙(복제 분기점)에서 시작된다는 점, DNA 중합효소는 DNA의 5`에서 3` 방향으로 이동한다는 점 때문에(DNA 복제가 5`에서 3`방향으로 복제 된다는 것을 감안하면), 텔로미어는 점차 부분적으로 짧아지게 된다.
선도 가닥에선 DNA 중합효소가 5`에서 3`로 쭉 진행하기 때문에 상호 보완적인 DNA 가닥을 만들 수 있다. 그러나 지연 가닥은 이와 같이 진행되지 않기 때문에 문제가 생긴다. 이 문제를 처리하기 위해서 프라이머 역할을 하는 짧은 RNA 조각 일부(RNA 프라이머)가 복제 분기점 앞 부분에 붙는다. DNA 중합효소는 그 부분에서부터 복제를 시작 시킬 수 있고, 복제 분기점에서 끝으로 이동한다(5`에서 3`방향으로). 그러나 지연 가닥에선 DNA 중합 효소가 연속적으로 끊김 없이 5`에서 3`로 이동할 수 없기 때문에 오카자키 절편이 만들어지게 된다. 더 많은 RNA 프라이머들이 DNA 가닥에 붙고 난 후에야 DNA 중합효소들이 계속해서 복제가 진행되도록 한다.
마지막 RNA 프라이머가 붙게되면 DNA 중합효소, RNA 뉴클레이스, DNA 연결효소가 RNA 조각 부분을 DNA로 변환시키고 오카자키 절편들의 사이를 메우기 위해 그 마지막 RNA 프라이머와 함께 온다. 그러나, RNA 부분(RNA 프라이머)을 DNA로 전환시키기 위해서는 RNA 프라이머의 앞부분에 또 다른 DNA 가닥(부분)이 있어야 한다. 그래야만 하는 이유가 말단의 RNA 프라이머가 제거된 후, 5` 쪽에는 dNTP를 중합할 3`–OH가 존재하지 않기 때문이다. 이것은 지연 가닥 모든 부분에서 항상 일어나는데 마지막 RNA 프라이머가 붙은 부분 끝에선 그 앞에 DNA부분이 없으므로 일어나지 않는다. 그러한 이유로, 이 경우에 해당하는 RNA 프라이머 부분은 복제된 DNA에서 RNA부분을 분해하는 효소에 의해 그냥 제거되어 버린다. 따라서 DNA 복제가 일어날 때 딸 세포의 지연가닥 5` 말단의 텔로미어는 지속적으로 점차 조금씩 사라진다.
그러나 몇몇 시험관에서 이루어진 연구들은 텔로미어가 산화스트레스(oxidative stress, 활성산소 참조)에 매우 민감하다는 것을 밝혔다. 또, 텔로미어의 결실(축소)에 있어 스트레스 매개 산화적인 DNA 손상(oxidative stress-mediated DNA damage)이 중요한 결정 요인이라는 증거도 있다. Long Island Breast Cancer Study Project (LIBCSP)에서 연구진은 가장 짧은 텔로미어를 가지고 있고 베타 카로틴, 비타민 C와 E 섭취를 적게하는 여성들이 그렇지 않은 여성들보다 유방암에 걸릴 위험이 조금 더 크다고 밝혔다. 이러한 결과들을 통해 텔로미어의 축소로 인한 암 유발 위험성이 DNA 손상, 산화 스트레스 같은 메커니즘들과 상호작용한다는 것을 유추해 볼 수 있다.
프라이머가 축소되는 현상은 노화, 나이와 연관된 질병들, 사망하는 것과 관련이 있다고 예측된다. 더 긴 텔로미어를 가진 생물이 짧은 텔로미어를 가지고 있는 생물보다 더 오래 살 수 있다는것은 Richard Cawthon가 2003년 밝혔지만, 짧은 텔로미어가 단순히 세포의 나이를 표시하는지 노화에 직접적으로 기여를 하는지는 밝혀진 바 없다. 노화가 진행되었다고 해서 모든 장기나 기관들의 세포에서 텔로미어가 일괄적으로 단축되지는 않는다. 또한 쥐의 텔로미어 길이(20~100 kb)가 우리의 것(5~15 kb)보다 김에도 불구하고 오래 살지 못하는 것을 보면 더욱 그러하다.
에이즈 바이러스(HIV)와의 연관성
에이즈 감염과 짧은 텔로미어의 연관성은 에이즈 바이러스가 세포, 생물학적 노화와 많은 나이와 관련된 질병의 영향에 간접적으로 영향을 주는 그 짧은 텔로미어에 영향을 준다고 제시했다. 실제로 에이즈 바이러스 감염군과 비감염군을 비교했을 때, 비감염군이 흡연율과 알코올 섭취량이 높았음에도 불구하고 바이러스 감염군의 텔로미어가 상당히 더 짧았다
흡연
한 연구에서 일생동안 담배를 한번도 피운적 없는 사람과 흡연 경험이 있는 사람들의 텔로미어 길이에 대하여 통계를 내었다. 흡연은 폐의 기능을 감소시켰고, 이것은 곧 긴 텔로미어의 축소를 야기했다. 한번도 흡연을 하지 않은 사람과 비교했을 때, 짧은 텔로미어를 가지는 흡연자들은 강제호기량(强制呼氣量)이 상당히 줄어들었다. 짧은 텔로미어를 가진 사람들은 흡연으로 인한 폐 기능 감소 현상이 가속화되었다는 것이다.
폐 이식 수술
폐 이식 수술에서 기증자와 환자 텔로미어 길이의 단면적인 특성은 이전에는 알려지지 않았다. 나중에야 획득되고 태어날 때부터 타고나는 텔로미어 축소 현상은 COPD(만성 폐색성 폐질환)와 IPF(특발성 폐섬유증)를 포함한 말기 폐병에서 언급되었기 때문에, 한 연구진은 폐를 이식 받은 환자가 기증자 보다 더 짧은 텔로미어를 가질것이라고 가설을 세웠다.
108명을 대상으로 조사한 결과, 이식 수술을 받은 환자들의 텔로미어 길이가 기증자들에 비하여 상당히 짧았다. 이러한 현상은 기증자와 환자의 나이가 같을 때에도 발견되었다. 환자들의 텔로미어 길이 감소는 선천적인 현상이나 가속화된 축소 때문일 것이다. 그러나 장기 이식 이후의 기증자와 환자의 변화된 텔로미어 길이 역할은 밝혀지지 않았다.
연장

제한된 세포 분열 현상은 Leonard Hayflick에 의하여 발견되었고, 이를 Hayflick limit(헤이플릭 한계, 배양기 속에서 세포가 생존하는 한계)라 일컫는다. 헤이플릭 한계 발견을 시발점으로, 텔로미어가 축소되는 현상을 발 묶는 아이디어는 후에 Michael D. West가 창립한 Geron Corporation의 연구진이 발견했다.텔로머레이스(텔로미어 신장 효소) 촉매 물질을 복제하여 특정 조건에서의 텔로머레이스 작용이 텔로미어 축소를 방지하여 인간의 세포를 불멸로 만들 수 있는지 확인하는 테스트를 했다. 1998년에 "텔로머레이스가 세포 수명을 연장시킬수 있다"라는 결과는 공개적으로 발표 되었고, 지금은 인체 세포를 불멸화 시킬 수 있을 것으로 기대된다.
텔로머레이스를 활성화시킴으로써 텔로미어 축소를 역으로 작용시켜 노화가 정상보다 느리게 진행되도록 할 수 있다는 것은 분명해지고 있다. 이는 헤이플릭 한계를 연장 시키므로 인간의 수명은 그에 따라 늘어날 것이다. 또한 약물, 유전자 치료, 무기력함이나 동면과 같이 물질 대사를 억제하는 이 세 가지 방법을 통해 텔로미어 축소를 반전시킬 수 있는 것으로 제시되었다. 아직까지 인체에선 앞서 제시된 방법들이 효과가 있다고 직접적으로 밝혀진 바는 없으나, 텔로미어 축소가 역으로 작용하는 동면 상태에서 노화가 억제된다는 것(Turbill, et al. 2012 & 2013)과 동면하는 것이 수명을 연장시킨다는 것(Lyman et al. 1981)은 입증되었다. 또 실험용 쥐와예쁜꼬마선충을 이용해 텔로미어 연장이 그 개체의 노화 신호들을 역으로 작용시킨 실험을 해냈다. 그러나 텔로머레이서가 작용하는 긴 텔로미어는 암세포가 될 확률이 높다고 가설이 세워졌다(e.g. Weinstein and Ciszek, 2002). 또, 그 역으로 긴 텔로미어는 암으로부터 보호를 하기도 하는데 짧은 텔로미어들은 암과 밀접하기 때문이다. 그리고 긴 텔로미어가 에너지 소비량을 증대시킬 수 있다고 제시되었다.
텔로미어를 연장시키는 기술은 생물 의학 치료 기술에 사용될 수 있는 충분히 많은 양의 건강하고, 암화되지 않은 포유류의 세포들을 배양하는 것을 가능하게 하기 때문에 조직공학에 도움이 될 것으로 전망된다.
장수(長壽)한 바닷새에 관한 두 연구는 텔로미어의 역할이 아직까지 완벽하게 밝혀지지 않았다는 것을 보여준다. 2003년, 과학자들은 바다제비(Leach's storm-petrel, Oceanodroma leucorhoa)의 텔로미어가 생활 연령에 따라 길어지는 현상을 관찰했다. 이러한 텔로미어 길이의 변화는 처음으로 관찰되었다. 그리고 2006년, Juola 외 몇명이 또 다른, 수명이 긴 바닷새 종인 큰군함조의 텔로미어 길이는 대략 최소한 40년 동안(전 생을 걸쳐) 지속적으로 줄어들었으나, 나이가 듦에 따라 그 속도가 확연하게 줄어들고 이러한 텔로미어 길이의 감소율은 각 새 개체에 따라 매우 다양했다. 연구진은 이 종(군함새와 그의 친척종들)에서 텔로미어의 길이는 한 새의 나이를 판별하기에 충분하지 않은 것이라 결론내렸다. 처음 믿어졌던 것 보다도 텔로미어 길이의 습성은 훨씬 더 다양한 것으로 기대된다.
텔로미어의 길이와 수명에 대한 의문점
고메즈 외 연구진(Gomes et al)이 포유류 동물 텔로미어의 비교 생물학적 연구에서 서로 다른 포유류 종의 텔로미어 길이는 수명과 직접적이진 않다고 밝혔으며 수명에 대한 텔로미어 길이의 영향이 여전히 의문이라고 결론을 내렸다. 헤리스 외 연구진(Harris et al)은 "인간 텔로미어 길이는 인지적 능력과 신체 능력에 대한 노화의 중요한 생물지표다"라는 증거를 찾았다. 길리와 블랙번(Gilley and Blackburn)은 짚신벌레를 이용하여 세포 노쇠기가 텔로미어 축소에 의한 것인지를 알아보기 위해 실험했으나, 세포 노쇠기 동안 텔로미어는 줄어들지 않았다.
생활 습관과의 연관성
캘리포니아 대학에서 추진된 한 준비 조사는 초기 전립선 암 진단을 받은 35명을 대상으로 했고 그 중 10명의 생활 방식을 바꾸게 했다. 채식(과일, 채소, 특별한 화학 처리가 안된 곡물, 저지방, 정제된 당질), 적당한 운동(1주일에 6일 30분씩 걷기), 스트레스 감소시키기(요가 스트레칭, 호흡, 명상), 주간 그룹 지원이 그 방식들이다. 그 외 25명 참가자들과 비교했을 때, 생활 습관을 바꾼 그룹에서 대략 10%의 더 긴 텔로미어가 관찰되었다. 게다가 얼마나 그 요청에 응했는지(얼마나 열심히 자신의 생활 방식을 바꾸었는지)에 따라서도 더욱 상당한 차이가 있었다. 2014년, "건강을 위해 일어서라. 앉아서 하는 행동을 피하는 것은 당신의 텔로미어를 연장시킬 것이다."란 표제를 지닌 한 연구는 다소 모순되는 결과를 제시했는데, 다음과 같이 결론을 내렸다. "중재 그룹에서 텔로미어 길이의 변화와 운동하는데 보낸 시간 변화 사이에 부정적인 연관성이 있었다(rho=-0.39, p=0.07). 그런데, 중재 그룹에서 텔로미어 연장은 앉아있는 시간을 줄이는 것과 상당히 연관이 있었다(rho=-0.68, p=0.02)"
텔로미어와 암세포
암세포는 끊임없이 세포분열을 하기 위해서 텔로미어 DNA의 길이를 유지할 수 있는 메커니즘이 필요하다. 텔로미어의 길이를 연장하거나 유지하는 메커니즘은 세포 단위의 끊임없는 증식을 하기 위해서 꼭 필요한 과정이다. 텔로미어를 연장하기 위해 필요한 효소인 텔로머레이스는 종양의 90%에서 활성화되어 있다. 그렇기 때문에 암세포에서는 다른 체세포에 비해서 수명이 길어진다. 이를 통해 암세포에 있는 텔로머레이스의 기능을 억제하거나 암세포의 텔로미어 DNA를 제거하면, 암세포의 세포분열을 막을 수 있다.
p53이나 pRb와 같은 종양억제유전자가 존재하면 세포노화가 일어나고 그렇지 않으면 어느정도 노화가 진행된 체세포가 암세포로 변할 수 있다.
최근 실험및 면역학 교수이자 이 대학의 벡스터 줄기세포 생물학연구소(Baxter Laboratory for Stem Cell Biology) 이사인 헬렌 블라우(Helen Blau) 박사는 인간 텔로미어 길이를 1,000 뉴클레오타이드 길이만큼 늘이는 방법(TERT 시퀀스가 코딩된 mRNA를 이용하여)을 발견했다. 이는 사람 수명으로 수천년에 해당한다.
기타
2009년 노벨상 수상위는 텔로미어와 텔로머레이스의 연구로 잭 쇼스택 외 2명에게 노벨 생리학·의학상을 수여했다.
참고 문헌
- Ricki Lewis 외,《LIFE》,6th Ed.,Mc Graw Hill(2009)
- 오상진, 노화의 생물학-분자생물학적 원인에 대하여(2015)
- Elizabeth Auld 외, HIV Infection Is Associated with Shortened Telomere Length in Ugandans with Suspected Tuberculosis(2016), University of North Carolina at Chapel Hill
- Gender and telomere length: systematic review and meta-analysis
- Sex differences in telomeres and lifespan.
- Associations Between Smoking, Telomere Length And Lung Function Decline: Findings From A Population-Based Longitudinal Study(2016)
| 역사 |  |
||||
|---|---|---|---|---|---|
| 하위 분야 | |||||
| 생물학적 개념 | |||||
| 일반 개념 | |||||
| 기본 기술 및 도구 |
|
||||
| 응용 | |||||
| 학제 간 분야 | |||||
| 목록 | |||||