
면역 글로불린 종류 변환(Immunoglobulin class switching, isotype switching, isotypic commutation 또는 class-switch recombination(CSR))은 B세포의 면역 글로불린 생산을 한종류에서 다른 종류로 바꾸는 생물학적 메커니즘이다. 예로는 아이소타입(isotype) IgM에서 isotype IgG로의 변환이 있다. 이 과정동안, 항체의 중쇄(heavy chain)의 불변부위는 변화하며, 중쇄의 가변부(variable region)는 그대로 남는다 (용어 '가변'과 '불변'은 다른 항원결정부(epitope)를 목표로 하는 항체의 변화 또는 변화없음을 일컫는다). 이 가변부(variable region)은 변하지 않기 때문에, 종류 변환은 항원 특이성(specificity)에 영향을 주지는 않는다. 대신, 항체는 동일 항원에 대한 친화도를 유지 하지만 다른 효과기(effector) 분자와 작용할 수 있다.
메커니즘
클래스의 전환은 성숙 B세포의 활성 이후에 일어난다. 이는 세포막의 항체분자 (또는 B세포 수용체(BCR))를 통해 일어나는데, 미성숙 B세포에서 만들어진 항체가 V(D)J 재조합 과정중에 항체는 가변부(variable domains)는 모두 동일하게 가지고 있지만, 중쇄(heavy chains)의 불변부(constant domains)는 다른것을 가지게 된다.
나이브(Naïve) 성숙 B 세포는 IgM과 IgD를 생성하는데, 이는 면역글로불린 자리(locus)의 첫 두 중쇄(heavy chain) 부분이다. 항원에 의해 활성화된 이후, 이 B 세포들은 증식한다. 만약 활성화된 B세포가 CD40이나 사이토카인 수용체를 통해 (둘 다 도움 T 세포에 의해 조절됨) 특정 신호 분자를 만나면, 이 B세포는 항체 종류 변환을 통해 IgG, IgA 또는 IgE 항체를 생성한다. 종류 변환 중에, 항체 중쇄의 불변부위는 변화하지만 가변부위는 변하지 않기 때문에, 항원에 대한 특이성은 변하지 않는다. 이러한 특징은 같은 활성화된 B 세포로부터 나온 다른 딸 세포들이 다른 아이소타입(isotype) 또는 서브타입(subtype) (예: IgG1, IgG2 등) 항체를 만드는 것을 가능하게 한다.
중쇄(heavy chain) 엑손(exons)의 순서는 다음과 같다.
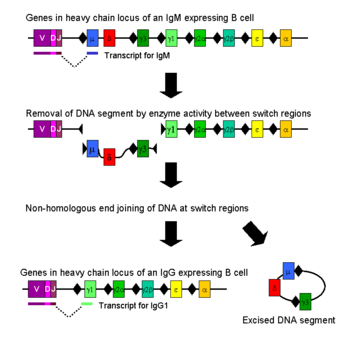
종류 전환은 종류 전환 재조합(class switch recombination(CSR))이라는 메커니즘에 의해 발생한다. 종류 전환은 생물학적인 메커니즘으로 활성화된 B세포에 의해 만들어지는 항체의 종류가 종류변환(isotype or class switching)이라는 과정동안 종류가 변하는 것이다. CSR 중에, 항체 중쇄 부위는 염색체로부터 제거되고, 삭제된 부분을 둘러싸는 유전자 조각은 항체 기능을 유지하기 위해 재결합한다. 이것이 다른 아이소타입(isotype)의 항체를 만들게 된다. 이중가닥의 손상부분은 DNA의 보존된 뉴클레오티드 모티프(nucleotide motifs)부위, 스위치 (S) 지역에서 만들어진다. 이는 항체 중쇄의 불변부를 인코딩 하는 유전자 상류에 있다. 이는 δ-체인을 제외하고는, 모든 중쇄의 불변부 주위에서 일어난다. DNA는 일련의 효소들의 활성에 의해 두 선택된 S 지역에 금이 가고 손상된다. 이러한 효소로는 AID(Activation-Induced(Cytidine) Deaminase), 우라실 DNA glycosylase와 AP(apyrimidic/apurinic)-endonuclease가 있다. S지역에 끼어든 DNA는 결과적으로 염색체에서 제거되며, 원치않은 μ 또는 δ 중쇄 불변부 엑손(exon)을 제거하며 이는 γ, α 또는 ε 불변부 유전자 조각이 대체하게끔 한다. DNA 끝 부위는 NHEJ(non-homologous end joining)라는 과정을 통해 재결합하여 가변부 엑손을 원하는 항체 중쇄의 하류 불변부 도메인 엑손과 연결한다. 동일하지 않은(non-homologous) 말단 접합이 없을 때, 자유로운 DNA 끝은 미세동일부(microhomology) 쪽으로 결합하는 다른 방법으로 재결합 할 수도 있다. μ 와 δ 유전자를 제외하고, 어떤 경우에도 한 개의 B세포는 한가지 종류의 항체만을 발현한다. 종류 전환 재조합이 대부분 삭제의 과정이지만, 염색체를 재배열하는 것은 발생할 수 있으며 (Ig 종류에 따라 10에서 20%), 이는 면역글로불린 중쇄 부위 양쪽에서 섞이는 염색체간 전위로써 발생할 수 있다.
종류 변환에 연관된 사이토카인
T 세포 사이토카인(cytokines)은 종류변환을 조절함이 쥐(표 1)와 인간(표 2)에서 확인됐다. 이러한 사이토카인(cytokines)은 IgM 생성을 억제할 수도 있다.
종류 변환에 관련된 유전자 조절 시퀀스(gene regulatory sequences)
목표 S 지역의 고도로 반복되는 구조에 덧붙여, 종류 변환의 과정은 S 지역이 처음 전사되고 (인트론 내에 있는) 면역글로불린 중쇄부위 전사물이 이어지는 과정이 필요하다. 크로마틴 리모델링, 전사와 AID 접근성, 손상된 S 지역의 접합(synapsis)은 3' 조절지역(3'RR)이라는 큰 슈퍼인핸서의 통제하에 있다. 이것은 Calpha이라는 먼 하류(distal downstream)에 위치한다. 어떠한 경우에는, 3'RR 슈퍼인핸서가 AID에 의해 표적이 되어 DNA 파괴와 Sµ와 접합을 겪을 수 있다. 이것은 면역글로불린 중쇄부 로커스(locus)를 지우며, 로커스 자살 재조합(locus suicide recombination, LSR)이다.
| T 세포 | Cytokines | 면역 글로불린스 클래스 | |||||
|---|---|---|---|---|---|---|---|
| IgG1 | IgG2a | IgG2b | IgG3 | IgG4 | IgE | IgA | |
| Th2 | IL-4 | ↑ | ↓ | ↓ | ↓ | ↓ | ↑ |
| IL-5 | ↑ | ||||||
| Th1 | IFNy | ↓ | ↑ | ↓ | ↑ | ↓ | ↓ |
| Treg | TGFß | ↑ | ↓ | ↑ | |||
| IL-10 | ↑ | ||||||
| T 세포 | Cytokines | 면역 글로불린스 클래스 | |||||
|---|---|---|---|---|---|---|---|
| IgG1 | IgG2 | IgG3 | IgG4 | IgA | IgE | ||
| Th2 | IL-4 | ↑ | ↓ | ↑ | ↑ | ||
| IL-5 | ↑ | ||||||
| Th1 | IFNy | ↓ | ↑ | ↓ | |||
| Treg | TGFß | ↓ | ↑ | ||||
| IL-10 | ↑ | ↑ | |||||
외부 링크
- 의학주제표목 (MeSH)의 Immunoglobulin+class+switching