
| 생물학 관련 문서 |
| 진화 |
|---|
 |
| 진화 과정 |
| 연구 및 역사 |
| 진화 이론의 주요 분야 |
| 주요 학자 |
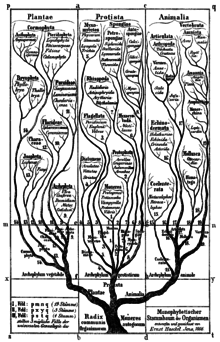
계통학(系統學, phylogenetics)은 여러 생물의 진화를 연구하는 학문이다. 분류계통학, 계통발생학이라고 하기도 한다.
이러한 관계는 DNA 서열, 단백질 아미노산 서열 또는 형태와 같은 관찰된 유전 특성을 종종 이러한 특성의 특정 진화 모델에서 평가하는 계통발생적 추론 방법에 의해 가정된다. 이러한 분석의 결과는 계통 발생(계통 발생 수라고도 함)으로, 집단의 진화 역사를 반영하는 관계에 대한 도식적 가설이다.
분류학은 생물의 식별, 명명 및 분류에 관한 학문이다. 분류는 이제 일반적으로 계통 발생 데이터를 기반으로 하며 많은 체계학자들은 단일 계통 분류군만 명명된 그룹으로 인식되어야 한다고 주장한다. 분류가 추론된 진화론적 역사에 의존하는 정도는 분류학파에 따라 다르다. 계통 발생 체계(cladistics)는 공유된 파생 특성(synapomorphies)을 기반으로 그룹만 인식하여 분류에 계통 발생을 반영하려고 한다. 진화적 분류는 분기 패턴과 차이점의 정도를 모두 고려하여 둘 사이의 절충안을 찾으려고 한다.
계통수
정의
계통수의 끝 부분은 살아있는 분류군이나 화석이 될 수 있으며 진화 혈통에서 끝 또는 현재를 나타낸다. 계통수는 뿌리를 내리거나 뿌리를 내리지 않을 수 있다. 뿌리가 있는 계통수의 뿌리는 가상의 공통 조상 또는 공통 혈통을 나타낸다. 뿌리가 없는 계통수는 조상 계보에 대해 가정하지 않으며 해당 분류군의 기원이나 뿌리 또는 추론된 진화적 변형의 방향을 나타내지 않는다.
분류군 사이의 계통 발생 패턴을 추론하기 위한 적절한 사용 외에도 계통 발생 분석은 종종 유전자 사본 또는 개별 생명체 간의 관계를 나타내는 데 사용된다. 이러한 사용은 생물다양성, 진화, 생태학 및 게놈을 이해하는 데 핵심이 되었다. 2021년 2월, 과학자들은 처음으로 동물의 유골(이 경우에는 매머드, 백만 년 이상 된 매머드)의 DNA 시퀀싱을 보고했으며, 이는 현재까지 시퀀싱된 가장 오래된 DNA이다.
계통수 추론
계통발생적 추론의 일반적인 방법에는 최적성 기준과 간결성, 최대 가능성 (ML) 및 MCMC 기반 베이지안 추론 방법을 구현하는 계산적 접근이 포함된다. 이 모든 것은 관찰된 진화를 설명하는 수학적 모델에 의존한다.
20세기 중반에 널리 사용된 페네틱스는 형태학의 전반적인 유사성 또는 유사한 관찰 가능한 특성(즉, 표현형 또는 DNA의 전체 유사성. DNA 서열이 아님)의 전반적인 유사성을 기반으로 계통수를 구성하기 위해 거리 매트릭스 기반 방법을 사용했다. 이는 종종 계통발생적 관계를 근사화하는 것으로 가정되었다.
1950년 이전에는 계통발생적 추론이 일반적으로 개연성을 가지는 모델로 제시되었다. 그러나 이 방법은 종종 모호하고 대안 가설을 평가하기 위한 명확한 기준이 없다.
현재는 NCBI를 비롯한 여러 사이트에 있는 미리 시퀀싱된 유전적 정보를 통해 누구나 쉽게 계통수를 추론할 수 있다.
계통군
계통학에서는 생물의 분류군을 분지학적으로 나눈다. 공통 조상과 그 자손을 모두 포함한 분류군은 단계통군이다. 파충류를 분류할 때 파충류로부터 진화한 새를 제외하는 경우가 있는데, 이와 같이 공통 조상은 포함했으나 모든 자손을 포함하지는 않은 분류군을 측계통군이라 한다. 온혈동물처럼 구성원들의 공통 조상을 포함하지 않은 분류군은 다계통군이라 부른다.
발전 과정
헤켈의 요약 이론
19세기 후반에 에른스트 헤켈(Ernst Haeckel)의 요약 이론 또는 생물유전학적 기본법칙이 널리 받아들여졌다. 이는 "개체 발생이 계통 발생을 반복한다는 것이다. 즉, 세균에서 성체까지의 일생 동안 단일 유기체의 발달은 그것이 속한 종의 연속 조상의 성체 단계를 연속적으로 반영한다. 그러나 이 이론은 오랫동안 거부되었고, 대신 개체 발생의 진화가 받아들여졌다. 헤켈이 생각했던 것처럼 종의 계통 발생 역사는 개체 발생에서 직접 읽을 수 없지만, 개체 발생의 특성은 계통 발생 분석을 위한 데이터로 사용될 수 있다. 분지학적으로 더 밀접하게 관련된 두 종이 가진 특성은 그들의 배아도 공유하는 특성이다.
중심 내용의 전개
- 14세기, lex parsimoniae (parsimony 원리), William of Ockam , 영국 철학자, 신학자, 그리고 Franciscan 수사, 그러나 그 아이디어는 실제로 아리스토텔레스로 거슬러 올라가 , 선구적인 개념
- 1763, 베이지안 확률, Thomas Bayes 목사, 선구자 개념
- 18세기, 피에르 시몽(Marquis de Laplace), 아마도 최초의 ML(최대 가능성) 사용, 전구체 개념
- 1809, 진화론, 철학자 동물학 , Jean-Baptiste de Lamarck , 선구적 개념, 볼테르, 데카르트, 라이프니츠가 17세기와 18세기에 예고했으며, 라이프니츠는 관찰된 간격을 설명하기 위해 진화적 변화를 제안하기까지 하여 많은 종들이 진화했다고 제안했다. 멸종되고 다른 종은 변형되었으며 공통 특성을 공유하는 다른 종은 한 번에 하나의 인종이었을 수 있으며 BC 6세기의 Anaximander 와 BC 5세기의 원자론자들과 같은 일부 초기 그리스 철학자들도 예고했다. 제안된 기본 진화 이론
- 1837년, 다윈의 노트는 진화적 나무를 보여줍니다
- 1843년, 상동 성과 유추의 구별 (후자는 현재 동종형성 이라고 함 ), Richard Owen, 선구자 개념
- 1858년, 고생물학자 하인리히 게오르크 브론(1800–1862)은 오래된 종의 멸종 이후에 새로운 유사한 종의 고생물학적 "도착"을 설명하는 가상의 나무를 발표했다. Bronn은 그러한 현상을 담당하는 메커니즘, 전구체 개념을 제안하지 않았다.
- 1858년, 진화론의 정교화, Darwin과 Wallace, 또한 다음 해 Darwin의 Origin of Species, 전구체 개념
- 1866년, Ernst Haeckel , 처음으로 계통발생에 기반한 진화수, 전구체 개념 발표
- 1893, 특성 상태 비가역성의 돌로 법칙, 전구체 개념
- 1912년, ML은 Ronald Fisher에 의해 권장, 분석 및 대중화됨, 전구체 개념
- 1921년, Tillyard는 "계통 발생"이라는 용어를 사용하고 그의 분류 시스템에서 고대 문자와 특수 문자를 구별하였다.
- 1940, Lucien Cuénot이 만든 "clade"라는 용어
- 1949, Jackknife 리샘플링 , Maurice Quenouille(Mahalanobis에 의해 '46년에 예고되고 Tukey에 의해 '58년에 확장됨), 전구체 개념
- 1950, Willi Hennig의 고전적 형식화
- 1952, William Wagner의 그라운드플랜 발산법
- 1953, "cladogenesis"가 만들어짐
- 1960, Cain과 Harrison이 만든 "cladistic"
- 1963년, 계통 발생학에 대해 ML(최대 가능성)을 사용하려는 첫 번째 시도, Edwards 및 Cavalli-Sforza
- 1965년
- Camin-Sokal parsimony, 최초의 parsimony(최적화) 기준 및 cladistic analysis를 위한 최초의 컴퓨터 프로그램/알고리즘, Camin과 Sokal
- Clique 분석이라고도 하는 문자 호환성 방법은 Camin과 Sokal(loc. cit.)과 EO Wilson 의해 독립적으로 도입되었다.
- 1966년
- Hennig의 영어 번역
- "cladistics" 및 "cladogram" 생성(Webster's, loc. cit.)
- 1969년
- 동적 및 연속 가중치, James Farris
- 바그너의 간결함, 클루게와 패리스
- CI(일관성 지수), Kluge 및 Farris
- 클리크 분석을 위한 쌍별 호환성 도입, Le Quesne
- 1970, Farris에 의해 일반화된 Wagner 간결함
- 1971년
- ML을 계통발생학(단백질 서열용)에 최초로 성공적으로 적용, Neyman
- 피치 패시모니, 피치
- NNI(최근접 이웃 교환)는 Robinson 과 Moore et al.에 의해 독립적으로 개발된 첫 번째 분기 교환 검색 전략이다.
- ME(최소 진화), Kidd 및 Sgaramella-Zonta (이 방법이 쌍별 거리 방법인지 또는 Edwards와 Cavalli-Sforza가 ML을 "최소 진화"라고 부르기 때문에 ML과 관련이 있는지 명확하지 않음)
- 1972, Adams 합의, Adams
- 1976, 계급에 대한 접두사 시스템, Farris
- 1977, 돌로 파시모니, 패리스
- 1979년
- 넬슨 합의, 넬슨
- MAST(최대 동의 하위 트리)((GAS)최대 동의 하위 트리), 합의 방법, Gordon
- bootstrap, Bradley Efron, 선구자 개념
- 1980, PHYLIP, 계통 발생 분석을 위한 최초의 소프트웨어 패키지, Felsenstein
- 1981년
- 다수의 합의, Margush 및 MacMorris
- 엄격한 합의, Sokal 및 Rohlf
- 최초의 계산 효율적인 ML 알고리즘, Felsenstein
- 1982년
- PHYSIS, Mikevich 및 Farris
- 브랜치 앤 바운드, 헨디와 페니
- 1985년
- 결합된 표현형 및 유전형 증거에 기초한 진핵생물의 첫 번째 분지 분석 Diana Lipscomb
- Cladistics의 창간호
- 부트스트랩의 첫 번째 계통 발생학적 응용, Felsenstein
- 잭나이프의 첫 번째 계통발생학적 응용, Scott Lanyon
- 1986, MacClade, Maddison 및 Maddison
- 1987, 이웃 결합 방법 Saitou와 Nei
- 1988, Hennig86(버전 1.5), 패리스
- Bremer 지원(쇠퇴 지수), Bremer
- 1989년
- RI(보존 지수), RCI(재배율 일관성 지수), Farris
- HER(동형 성형 초과 비율), Archie
- 1990년
- 결합 가능한 구성 요소(준 엄격) 합의, Bremer
- SPR(서브트리 가지치기 및 재접합), TBR(나무 이분 및 재연결), Swofford 및 Olsen
- 1991년
- DDI(데이터 결정성 지수), Goloboff
- 표현형 증거만을 기반으로 한 최초의 진핵생물 분류학적 분석, Lipscomb
- 1993, 묵시적 가중치 Goloboff
- 1994, 감소된 합의: 뿌리 나무에 대한 RCC(감소된 분지 합의), Wilkinson
- 1995, 루팅되지 않은 트리에 대한 감소된 합의 RPC(감소된 파티션 합의), Wilkinson
- 1996년 Li, Mau, 및 Rannala와 Yang 의해 독립적으로 개발된 BI(Bayesian Inference)에 대한 최초의 작업 방법 과 모두 MCMC(Markov chain-Monte Carlo)를 사용하여 개발됨
- 1998, TNT(신기술을 사용한 나무 분석), Goloboff, Farris 및 Nixon
- 1999, Winclada, 닉슨
- 2003, 대칭 재표본, Goloboff
- 2004,2005, 대칭성 메트릭(Kolmogorov 복잡성에 대한 근사값 사용) 또는 NCD(정규화된 압축 거리), Li et al., Cilibrasi 및 Vitanyi.
외부 생물학
계통 발생 도구 및 표현(계통수와 유전적 네트워크)은 양적 비교 언어학 분야에서 언어의 진화를 연구하는 데에도 적용될 수 있다.