
| 히스톤 | |
|---|---|
 | |
| 식별자 | |
| PDB | 1AOI |
히스톤(Histone)은 생물학에서 염색질(chromatin)을 구성하는 기본단위인 뉴클레오솜(nucleosome)의 중심 단백질이다. 이들은 DNA 사슬이 감기는 실패 역할을 해서 DNA의 응축을 도우며, 유전자 발현조절에 중요한 역할을 한다.
분류
6종류의 주요한 히스톤이 알려져 있다.
히스톤 H2A, H2B, H3, H4는 각각 2개씩 모여서 8합체인 이른바 핵심 히스톤(core histone)을 구성한다. 히스톤은 뉴클레오솜 핵 부분에 존재하며 146 염기쌍의 DNA를 감으며, DNA는 하나의 히스톤을 왼손방향으로 1.65 회 정도 감는다. 연결히스톤 H1은 뉴클레오솜 DNA의 감기지 않은 부분에 달라붙어서 DNA를 고정하며, 더 높은 차원의 구조를 형성할 수 있도록 한다. 이 구조의 가장 기본적인 형태는 10nm의 구슬이 실에 꿰어져 있는 것으로 묘사된다. 이 구조는 뉴클레오좀솜이에 50염기쌍 정도의 DNA가 있다는 사실을 포함하고 있다. 이렇게 생성된 히스톤과 DNA의 결합체를 염색질이라고 불린다. 더 높은 구조는 비정상적인 지그재그 구조를 형성하는 30 nm 섬유와 100 nm 섬유구조를 포함하며, 정상적인 세포에서 발견된다. 체세포분열과 생식세포분열 동안에, 응축된 염색질들은 뉴클레오좀과솜른 조절단백질 사이의 상호작용을 통해서 조합된다.
구조
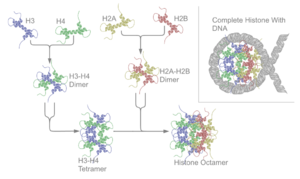
뉴클레오솜핵은 H2A-H2B 이합체와 H3-H4 사합체가 모여서 생긴 팔합체의 4차구조를 가지며, 그 대칭성은 대칭면을 1개 갖는 C2의 점군 대칭성을 갖는다.. H2A-H2B 이합체와 H3-H4 사합체는 슈도다이아드 대칭성을 갖는다. 4종류의 중심 히스톤들(H2A, H2B, H3, H4)들은 상대적으로 비슷한 구조를 가지며, 진화과정에서 잘 변하지 않는다, 그들은 긴 꼬리의 말단의 같은 아미노산 서열을 가지며, 이는 전사후변이에 중요한 역할을 한다.
히스톤은 DNA와 아래의 5종류의 상호작용을 한다.
- 전체적으로 음전하를 띠는 DNA의 인산골격과 전체적으로 중심히스톤의 양전하 사이의 H2B, H3, H4 의 알파나선에 의한 나선 쌍극자
- DNA 골격과 히스톤의 중심사슬의 아민그룹 사이의 수소결합
- 히스톤과 DNA의 디옥시리보스 사이의 비극성 상호작용
- 염기성 아미노산(특히 리신과 알기닌)의 측쇄와 DNA의 인산기의 산소 사이의 염 연결과 수소결합.
- H3와 H2B 의 N말단 꼬리가 두개의 DNA 분자의 작은 홈으로 파고들면서 생기는 비특이성 삽입
히스톤의 높은 염기성은 DNA와 히스톤의 반응을 도울 뿐만 아니라 히스톤의 수용성에도 기여한다.
히스톤은 N 말단 꼬리 뿐만 아니라, 구형 영역에서도 효소에 의한 번역후 변이의 주체이다. 이런 변경은 메틸화, 시트룰린화, 인산화, SUMO화, 유비퀴틴화 ADP-리보스화를 포함한다. 이들은 그들이 작용된 유전자의 발현을 조절한다.
간기 동안, 비활성 유전자가 히스톤과 강하게 상호작용하고 있는 것과는 달리, 일반적으로 활동적인 유전자는 주변에 더 적은 히스톤을 갖고 있다. 이는 히스톤이 유전자 조정에 매우 중요하므로, 위험한 돌연변이가 쉽게 도태되므로 히스톤의 구조가 진화적으로 잘 보존된다는 것을 나타낸다.
기능
DNA 가닥 응축
히스톤은 DNA 가닥이 감기는 실패와 같은 역할을 한다. 이는 진핵생물의 거대한 유전자가 세포 핵으로 들어가기 위해 필수적인 응축을 할 수 있도록 한다. 응축이 종결된 분자는 500,000배나 짧아진다.
염색질 조절에서의 히스톤 변경
히스톤은 DNA와 핵단백질 사이의 상호작용을 바꾸는 번역의 개시(on)와 종결(off)의 변동과 함께 후생유전학(epigenetics)적 변경을 겪을수있다. H3, H4 히스톤은 뉴클레오솜서 튀어나와 있는 몇몇 장소에서 공유결합적으로 변경될 수 있으며 이를 위한 비교적 긴 꼬리를 갖고 있다. 꼬리에서의 반응은 메틸화, 아세틸화, 인산화, 유비퀴틴화 등을 포함한다. 히스톤의 중심(H2A, H3히스톤) 또한 변경될 수 있다. 이들 조합의 변경은 일명 히스톤 코드라는 코드를 구성하는 것으로 생각된다. 히스톤 변경은 유전자 조정, DNA 수리, 체세포분열과 같은 다양한 생물학적 기작에 관여한다.
히스톤 변경의 일반적인 명명법은 다음과 같다.
- 히스톤의 이름(예 : H3)
- 아미노산의 1글자 축약형(예 : 리신->K)과 단백질에서의 아미노산의 위치
- 변경의 종류 (Me : 메틸화, P : 인산화 , Ac : 아세틸화, Ub : 유비퀴틴화)
따라서 H3K4Me는 H3 히스톤의 N말단으로부터의 4번째 리신이 메틸화되었음을 의미한다.
전사 조정에서의 히스톤 변경에 대한 더 자세한 예는 염색질 구조에 따른 RNA 합성효소의 조절에서 볼 수 있다.
역사
히스톤은 1884년, 알브레히트 코셀에 의해 발견되었다. "히스톤"이라는 단어는 19세기 후반부터 사용되었으며, 어원은 명확치 않으나 아마도 그리스어 histanai 나, histos에서 유래되었을 독일어 "Histon"의 형태로 최초로 사용되었다. 1990년대 초반까지, 히스톤은 핵 DNA를 응축하는 물질일 것이라고만 생각되었으나, 1990년 초반부터 히스톤이 DNA의 발현을 조절한다는 것이 발견되었다.
종 사이의 보존성
히스톤은 진핵생물의 세포와 진핵고균(Euryarchaea)라 불리는 고균의 핵에서 발견된다. 그러나 세균에서는 발견되지 않는다. 고균의 히스톤은 아마도 진핵세포의 히스톤의 진화적 전구체와 매우 유사할 것이다. 히스톤 단백질은 핵의 생물학적 역할에서 매우 중요하기 때문에 진핵생물 내에서 잘 보존되어 있다
중심히스톤은 매우 잘 보존된 단백질이다. 그것은 다양한 종들의 히스톤 단백질사이에서 매우 적은 아미노산 서열 변화만이 있음을 의미한다. 연결히스톤은 종종 한 종에서 하나 이상의 형태가 있으며, 이는 중심히스톤보다 덜 보존됨을 나타낸다.
몇종류의 주요 히스톤의 변종들이 존재한다. 이들 변종들은 원종과 유사한 아미노산 서열과 구조를 갖고 있지만, 그들과 구분되는 차이를 갖고 있으며 이들을 '소량 히스톤'이라 부른다. 이들 소량히스톤들은 염색질대사에서 고유한 역할을 수행한다. 예를 들어 히스톤 H3과 비슷한 CenpA는 염색체의 동원체부위에만 결합하는 히스톤이다. 히스톤 H2A의 변종인 H2A.Z는 활발하게 전사되는 유전자의 촉진부위(promoter)에 달라붙으며 또한 이형염색체의 형성에 수반된다. 또다른 H2A의 변종인 H2A.X는 DNA 이중사슬이 부서진 곳에만 달라붙어서 DNA 수리가 진행 중인 지역을 표시한다. 히스톤 H3.3는 활발하게 전사되는 유전자의 본체에 결합한다.
뉴클레오솜
DNA 가닥(string)에 감긴 8개의 코어 히스톤(core histones-2개의 H2A ,2개의 H2B,2개의 H3,2개의 H4)과 히스톤 꼬리들(tails) 그리고 H1 히스톤이 뉴클레오솜을 주요하게 구성한다. 이러한 뉴클레오솜들이 염색질(chromatin)의 기본단위가 된다.
 |
그래픽적인 예시 |
아세틸기(acetylation)에 의해서 히스톤꼬리변형(histone tail modification)이 일어남으로써 DNA가 풀리고 닫히며 이러한 과정에서 유전자 복제,RNA복사본,RNA 3염기서열 당 한개의 아미노산 생성이 결정되게되는 그래픽적 예시를 보여준다.
같이 보기
외부 링크
- Chromatin, Histones & Cathepsin; PMAP The Proteolysis Map-animation
- Nextbio Archived 2009년 3월 2일 - 웨이백 머신
- Chromatin Regulation Signaling Pathways