




a. 분열하지 않는 세포.
b. 세포 분열을 준비하는 세포핵.
c. 분열하는 세포와 방추섬유.
e. 분열 직후의 딸세포 2개(왼쪽과 오른쪽).
세포생물학에서 체세포분열(mitosis /maɪˈtoʊsɪs/; 유사분열)은 복제된 염색체가 두 개의 세포핵으로 분리되는 세포 주기의 한 부분이다. 세포 분열은 전체 염색체 수를 유지하면서 유전적으로 동일한 세포를 만든다. 따라서 체세포분열을 균등 분열(equational division)이라고도 한다. 일반적으로 사이기의 S기 이후에 체세포분열(핵의 분열)이 진행되고, 체세포분열의 말기 이후에는 세포질과 세포소기관, 세포막이 하나의 세포에서 2개의 새로운 세포로 거의 동일하게 나뉘는 세포질분열이 일어난다. 체세포분열 과정의 여러 단계를 함께 M기(체세포분열기)라고 정의한다.
체세포분열의 과정은 특정 세포의 활동에 따라 여러 단계로 나뉘는데, 각 단계를 순서대로 전기, 전중기, 중기, 후기, 말기라고 한다. 체세포분열 동안 염색체는 복제되어서 응축된 상태이고, 각 복제된 염색체는 방추사에 부착하여 분리되면서 세포의 양 끝으로 끌려간다. 체세포분열 말기에 형성된 2개의 딸세포 핵은 유전적으로 서로 동일하다. 세포의 나머지 부분은 세포질분열 과정을 거쳐 2개의 딸세포를 만든다. 체세포분열의 오류로 3개 이상의 딸세포를 만드는 경우를 세극 체세포분열이라고 하거나 다극 체세포분열이라고 한다. 체세포분열에서 나타나는 다른 오류는 어팝토시스(프로그래밍된 세포 죽음)를 유도하거나 돌연변이를 일으킨다. 일부 암도 이러한 돌연변이로부터 생긴다.
체세포분열은 진핵생물의 세포에서만 일어난다. 원핵생물의 세포는 핵이 없기 때문에 이분법이라는 다른 방법으로 분열한다. 체세포분열 방법은 생물에 따라 다양한데, 예를 들어서 동물 세포는 염색체가 분리되기 전에 핵막이 쪼개지는 "열린" 체세포분열을 하고, 진균 세포는 세포핵이 그대로 존재하는 상태에서 염색체가 나뉘는 "닫힌" 체세포분열을 한다. 대부분의 동물 세포는 체세포분열을 시작할 때 구형 형태를 취하기 위해 체세포분열 세포 라운딩(mitotic cell rounding)이라는 모양 변화를 겪는다. 대부분의 사람 세포는 체세포분열로 만들어지지만, 정자와 난자 같은 생식자는 예외적으로 감수분열을 통해 만들어진다.
발견
세포 분열은 18세기와 19세기에 처음으로 많이 묘사되었다. 1835년 독일의 식물학자 후고 폰 몰(Hugo von Mohl)은 녹조류인 Cladophora glomerata가 세포 분열을 통해 세포 증식이 시작되는 것을 묘사했다. 1838년에는 다른 독일의 식물학자 마티아스 야콥 슐라이덴(Matthias Jakob Schleiden)이 식물의 내부에서 나타나는 새로운 세포 형성이 세포 증식의 일반적인 법칙이라는 사실을 확인했지만, 나중에 로버트 레막(Robert Remak) 같은 과학자들에 의해 몰(Mohl) 모델이 지지되면서 이 견해는 받아들여지지 않았다.
동물 세포에서 체세포분열은 1873년 개구리, 토끼, 그리고 고양이의 각막 세포에서 발견됐고, 1875년 폴란드의 조직학자 바츠왑 마이젤(Waclaw Mayzel)이 처음으로 이를 묘사했다.
부츨리(Bütschli), 슈나이더(Schneider), 폴(Fol)도 체세포분열 과정을 발견했을 수 있다. 1873년 독일의 동물학자 오토 부츨리(:en:Otto BütschliOtto Bütschli)는 선형동물을 관찰한 데이터를 출판했다. 몇 년 뒤 부츨리는 그 관찰을 기반으로 체세포분열을 발견하고 묘사했다.
"체세포분열(mitosis)"이라는 용어는 그리스어로 날실을 뜻하는 μίτος (mitos)에서 유래하여 1882년 발터 플레밍(Walther Flemming)이 만들었다. 1878년 슐라이허(Schleicher)가 도입한 "핵분열"이라는 용어나, 아우구스트 바이스만(August Weismann])이 제안한 "균등 분열"이라는, 체세포분열 과정을 의미하는 다른 용어도 존재한다. 그러나, "체세포분열"이라는 용어는 일부 학자들이 핵분열과 세포질분열을 동시에 나타내는 넓은 의미로 사용한다. 현재는 "균등 분열"이라는 용어가 체세포분열을 닮은 감수분열 과정 중 하나인 감수 2분열을 나타낼 때 더 일반적으로 사용된다.
그리스어 어원에서 살펴봤듯이 "mitosis"는 날실 같은 방추섬유가 관여하는 세포 분열이다. 일본은 이 용어를 처음 번역할 때 실이 존재하는 분열이라는 뜻에서 "有絲分裂"이라고 한자로 번역했고, 한국은 일본의 영향을 받아 한자를 그대로 읽어서 "유사분열"이라고 사용해왔다. 이후 이 단어를 우리말로 바꾸는 과정에서 "체세포분열"이라는 용어로 굳어졌고, "mitosis"를 번역할 때에는 "체세포분열"로 번역하는 것이 권장된다. 그러나 이는 원래 용어의 어원을 살리지 못한 용어이므로, 더 적절한 우리말 용어가 만들어질 필요성이 있다.
단계
개요
체세포분열과 세포질분열을 하면 모세포의 지놈이 2개의 딸세포로 전달된다. 염색체에서 DNA는 세포 기능 유지에 필수적인 유전 정보를 저장하고 있는데, 한 생물의 전체 유전 정보를 지놈이라고 한다. 각 딸세포는 모세포와 유전적으로 동일해야 하기 때문에 모세포는 반드시 체세포분열을 하기 전에 염색체의 사본을 만들어야 한다. 이는 사이기의 S기 때 일어난다.염색체 복제가 일어나면 2개의 동일한 자매 염색분체가 생기며, 각 자매 염색분체는 중심절(동원체)에서 코히신(cohesin)이라는 단백질로 서로 묶여있게 된다.
체세포분열이 시작되면 염색체는 응축하여 광학 현미경으로 보이게 된다. 진핵생물에서 핵막은 DNA를 세포질로부터 격리시키는 역할을 하는데, 체세포분열이 진행되면 일부 진핵생물, 예를 들어 동물 세포는 핵막을 작은 소포들로 해체시킨다. 핵에서 라이보솜을 만드는 핵소체도 사라진다. 미세관(microtubule)은 세포의 반대편 끝으로 뻗어나가 중심절에 부착하고 염색체를 세포 중앙에 배열시킨다. 이후 미세관은 각 염색체의 자매 염색분체를 끌어당겨 복제된 염색체를 분리시킨다. 분리된 직후 자매 염색분체는 딸염색체라고 부른다. 세포가 늘어나면서 각 딸염색체는 세포의 반대편 끝을 향하여 당겨지고, 후기의 후반부에서 최대로 응축한다. 말기에는 새로운 핵막이 분리된 딸염색체 주변에서 형성되고, 딸염색체들은 풀려서 사이기 핵을 형성한다.
체세포분열이 진행될 때 일반적으로 후기가 진행되고 나면 세포는 세포질분열 과정을 겪는다. 동물 세포의 경우 만들어지고 있는 2개의 핵 사이로 세포막이 세포 안쪽으로 집혀 들어간다. 식물 세포의 경우 세포판이 2개의 핵 사이에서 형성된다. 세포질분열이 항상 일어나는 것은 아니며, 일부 다핵세포는 세포질분열 없이 체세포분열을 진행한다.

사이기(간기) (Interphase)
체세포분열기(M기)는 세포 주기에서 상대적으로 짧은 시기이다. 체세포분열기가 끝나면 세포 분열 과정을 준비하기 위해 세포는 훨씬 긴 사이기를 거친다. 사이기는 G1기, S기, G2기의 3가지 시기로 나뉜다. 이때 G는 간격을 의미하는 'gap'의 약자이고, S는 합성을 의미하는 'synthesis'의 약자이다. 사이기의 세 시기를 거치는 동안 세포는 단백질과 세포소기관을 만들면서 성장한다. 그러나 염색체는 S기에서만 복제된다. 정리하면, 세포는 G1기 때 성장하고, S기 때 염색체를 복제하면서 계속 성장하고, G2기 때 체세포분열을 준비하면서 성장하고, 최종적으로 M기 때 분열하면서 세포 주기를 새로 시작한다. 세포 주기의 이 모든 시기는 사이클린과 사이클린-의존성 카이네이스, 그리고 다른 세포 주기 단백질에 의해 정밀하게 조절된다. 어떤 시기에서 다른 시기로 넘어가는 일은 매우 엄격하게 진행되고, 각 시기 사이에는 세포가 세포 주기를 진행할 수 있게 해주는 신호인 "체크포인트"가 존재한다. 세포 주기를 일시적 또는 영구적으로 벗어나 분열하지 않는 세포는 G0기로 접어든다. 이는 세포가 너무 많아져서 밀도-의존성 억제가 나타나거나, 혹은 사람 심장근육 세포나 뉴런처럼 세포가 개체에서 특정 기능을 갖도록 분화했을 때 나타난다. 일부 G0기 세포는 다시 세포 주기를 진행시킬 수 있다.
사이기 때는 2개의 주요 과정을 통해 DNA 이중가닥 절단을 복구할 수 있다. 첫 번째 과정은 비상동 말단연결(non-homologous end joining; NHEJ)인데, 이 과정을 통해 사이기의 모든 시기에 잘린 DNA의 끝부분을 서로 연결시킬 수 있다. 두 번째 과정은 상동 재조합 복구(homologous recombinational repair; HRR)인데, 이 과정은 NHEJ보다 더 정확한 DNA 이중가닥 절단 복구 방법이다. HRR을 하려면 서로 인접한 상동 서열이 필요하기 때문에 DNA 복제가 이루어지고 있는 S기나 DNA 복제가 다 이루어진 G2기 때만 할 수 있다.
사이기는 세포가 체세포분열을 준비하도록 도우며, 체세포분열이 일어날지 말지 결정한다. 세포의 DNA가 손상받거나 중요한 시기를 완료하지 못하면 세포 주기를 중단시킨다. 이처럼 사이기는 체세포분열이 성공적으로 완료될지 말지를 결정하는 데 매우 중요한 시기이다. 사이기에 관여하는 주요 단백질들이 실수라도 하면 잠재적인 암세포를 만들 수도 있다. 현재 각 시기들이 어떻게 일어나는지 구체적으로 이해하기 위하여 더 많은 연구가 진행되고 있다.
체세포분열기(M기) (Mitotic (M) phase)
체세포분열 (Mitosis)

전전기(식물 세포) (Preprophase)
식물세포에서만 전기 이전에 전전기(preprophase)가 나타난다. 공포(액포)가 매우 큰 식물 세포에서는 체세포분열이 일어나기 전에 세포핵이 세포의 중앙으로 이동해야 한다. 격막형성질(phragmosome)이라는 세포질을 가로지르는 판 모양의 구조물은 세포를 2등분하면서 핵을 중앙으로 옮긴다. 또한, 미세관 고리와 액틴 필라멘트가 형성되면서 세포막 아래에 적도판을 둘러싸며 전전기 띠(preprophase band)가 형성된다. 이 띠는 세포가 최종적으로 나뉠 위치를 표시해준다. 꽃을 피우는 꽃식물(현화식물)과 같은 고등 식물의 세포는 중심립이 없는 대신에 미세관이 핵의 표면에서 방추체를 형성하며, 핵막이 분해된 이후에는 염색체 스스로가 방추체를 조직화한다. 전전기 띠는 핵막이 분해되고 전중기에 방추체가 형성되면서 사라진다.
전기 (Prophase)


사이기의 G2기 이후에 진행되는 전기 때는 염색체를 고도로 응축하고 방추체를 형성하면서 세포가 분열될 준비를 한다. 사이기 동안 핵 안에서의 유전 물질은 느슨하게 풀어져서 염색질을 이뤘다. 그러나 전기가 시작되면 염색질 섬유가 응축하여 뚜렷하게 구분되는 염색체가 만들어지고, 이때 고도로 응축된 염색체는 광학 현미경을 사용하여 높은 배율에서 관찰할 수 있다. 이 시기의 염색체는 길고 가는 실처럼 생긴 구조물을 형성한다. 각 염색체는 복제된 형태이기 때문에 2개의 염색분체를 가진다. 각 염색분체는 중심절에서 서로 붙어 있다.
전기 동안 유전자 전사는 멈추고 후기의 후반부에서 G1기가 시작하기 이전까지 전사는 일어나지 않는다.핵소체도 전기의 초반에 사라진다.
중심체는 체세포분열 전기 때 동물 세포의 핵 근처에 존재하는 구조물이다. 각 중심체는 한 쌍의 중심립을 가지며, 여러 단백질이 느슨하게 이를 둘러싸고 있다. 중심체는 세포의 미세관을 조직화하는 중심이다. 세포는 사이기 때 중심체를 복제하고, 세포 분열할 때 딸세포는 하나의 중심체를 물려받는다. 전기 때 존재하는 2개의 중심체는 튜뷸린을 중합하면서 미세관 방추장치가 형성되도록 한다. 이후 운동 단백질은 미세관을 따라 중심체를 밀면서 세포의 양끝으로 이동시킨다. 중심체가 미세관 조립을 조직화하기는 하지만, 식물은 중심체가 없기 때문에 이것이 방추장치를 형성하는 데 꼭 필요한 것은 아니다. 중심체가 없는 식물은 핵막에서 방추장치가 형성되며, 동물 세포의 체세포분열에도 필수적으로 필요한 것은 아니다.
전중기 (Prometaphase)
동물 세포에서 전중기가 시작되면, 핵 라민이 인산화되면서 핵막이 작은 소포들로 분해된다. 핵막이 분해되면서 미세관은 핵의 영역까지 뻗어 들어온다. 이런 과정을 열린 체세포분열이라고 하고, 이는 일부 다세포 생물에게서 일어난다. 진균이나 조류나 트리코모나드 같은 일부 원생생물은 핵 내부에서 방추체가 형성되거나, 미세관이 핵막을 뚫고 들어가는 닫힌 체세포분열을 한다.
전중기의 후반부에는 방추부착점 미세관은 염색체의 방추부착점을 찾아서 부착한다.방추부착점이란 전기의 후반부 때 염색체의 중심절(centromere) 부근에 형성되며, 미세관이 부착할 수 있도록 해주는 단백질 구조물이다. 참고로 '방추부착점(kinetochore)'과 '중심절(centromere)'은 모두 '동원체'라고 잘못 번역되어 오류를 야기하므로 '동원체'라는 표현을 지양할 것을 권장한다.
한쪽 중심체에서 뻗어 나오는 많은 수의 극성 미세관은 다른 쪽 중심체에서 뻗어 나오는 극성 미세관을 찾아 상호작용하면서 방추체를 형성한다. 방추부착점의 구조와 기능이 완벽하게 밝혀지진 않았지만, 이는 어떤 형태의 분자 모터를 가지고 있다. 미세관이 방추부착점에 연결되면, 모터가 활성화되면서 ATP 에너지를 사용하며 중심체 방향으로 미세관을 기어간다. 이 모터 활성은 미세관의 중합 및 탈중합과 짝지어져서 나중에 염색체를 구성하는 2개의 염색분체가 분리될 때 필요한 당기는 힘을 만들어준다.
중기 (Metaphase)


전중기 때 미세관이 방추부착점에 위치한 뒤 부착하면, 2개의 중심체는 세포의 양쪽 끝으로 염색체를 당기기 시작한다. 이렇게 팽팽한 상태가 되면 염색체는 2개의 중심체 사이에 존재하는 가상의 평면(세포의 가운데 평면)인 중기판(적도판)에 배열된다. 체세포분열이 끝날 때 염색체를 공평하게 분배하기 위해서 중기 체크포인트가 검문을 실시하는데, 중기 체크포인트는 방추부착점이 방추와 적절하게 부착되었는지 그리고 염색체가 중기판을 따라 배열되었는지 확인한다. 세포가 중기 체크포인트를 성공적으로 통과하면, 후기가 진행된다.
후기 (Anaphase)

후기 A 때는 자매 염색분체를 서로 연결하고 있는 코히신(cohesin)이라는 단백질이 잘리면서 2개의 동일한 딸염색체를 만든다. 방추부착점 미세관은 짧아지면서 분리된 딸염색체를 세포의 양쪽 끝으로 당긴다. 후기 B 때는 극성 미세관이 서로를 밀면서 세포가 양쪽으로 길어지게 한다. 후기의 후반부에는 염색체가 최대로 응축하면서 염색체 분리(chromosome segregation)가 일어나며 이후 말기 때 핵이 다시 형성되도록 해준다. 핵이 다시 형성된다. 대부분의 동물 세포에서 후기 A가 후기 B보다 먼저 일어나지만, 일부 척추동물의 알세포는 후기 B가 후기 A보다 먼저 일어난다.
말기 (Telophase)

말기는 전기와 전중기 때 일어났던 사건이 반대로 일어난다. 극성 미세관은 계속해서 길어지면서 세포를 더 길게 늘린다. 핵막이 분해되었다면, 새로운 모세포의 오래된 핵막이 쪼개졌던 막 소포를 이용해 핵막이 다시 생긴다. 새로운 막은 각 분리된 딸염색체 주변으로 형성되며(중심체는 핵막 밖에 위치한다), 핵소체가 다시 나타난다. 나누어진 각 염색체 세트는 각각의 새로운 핵막으로 둘러싸이게 되고, 응축되었던 염색체는 탈응축되어 풀린다. 이로써 체세포분열이 완료된다. 각 딸핵은 동일한 염색체 세트를 가진다. 생물에 따라서 이 시점에서 세포 분열은 또 일어날 수도 있고 안 일어날 수도 있다.
세포질분열 (Cytokinesis)


세포질분열은 체세포분열의 시기에 포함되지는 않지만 체세포분열기(M기)에는 마지막 시기로 포함되어 세포 분열을 완료시킨다. 동물 세포와 식물 세포 둘 다 세포 분열은 골지체에서 유래한 소포에 의해 일어나며, 골지체는 미세관을 따라 세포의 가운데로 이동한다. 동물 세포에서는 수축고리를 가지는 분열고랑(cleavage furrow)이 중기판이었던 부위에 집게처럼 만들어져 각 분리된 핵을 집어서 나눈다. 식물 세포에서는 골지체 구조가 합쳐져 격벽형성체(phragmoplast)의 중심에서 세포판을 형성하고, 이것이 발달해서 세포벽이 되어 2개의 딸핵을 분리시킨다. 격벽형성체는 일반적으로 고등 식물에서 나타나는 미세관 구조이고, 일부 녹조류는 해조체(phycoplast)를 이용하여 세포질분열 때 미세관을 배열한다. 각 딸세포는 모세포와 완전히 똑같은 사본을 가진다. 세포질분열이 끝나면 체세포분열기(M기)도 함께 끝난다.
여러 세포들은 체세포분열과 세포질분열이 각각 따로 일어나면서 하나의 세포가 여러 개의 핵을 가지기도 한다. 이런 현상은 진균이나 점균류, 다핵 조류에서 두드러지게 관찰할 수 있다. 심지어 일부 동물도 세포질분열과 체세포분열이 독립적으로 일어나기도 한다. 초파리의 배아 발생 때 나타나는 특정 단계가 대표적인 예시다.
기능
체세포분열의 기능과 중요성은 온전한 염색체 세트를 유지하는 데 있다. 각 딸세포는 모세포와 동일한 염색체 구성과 염색체 수를 전달받는다.
체세포분열로 나타나는 현상에는 다음과 같은 것들이 있다.
- 성장과 발생: 한 생물을 구성하는 세포의 수는 체세포분열을 통해 증가한다. 체세포분열을 통해 하나의 세포로부터 다세포로 이루어진 몸으로 발생할 수 있다.
- 세포 대체: 피부나 소화계 같은 몸의 일부분은 끊임없이 사라지고 새로운 세포로 대체된다. 새 세포는 체세포분열을 통해 형성되며, 대체될 세포와 똑같은 사본이다. 적혈구는 4개월이라는 짧은 수명을 지녔는데, 적혈구가 죽으면 체세포분열로 형성되는 새로운 적혈구로 대체된다.
- 재생: 일부 생물은 몸의 일부분을 재생시킬 수 있다. 새로운 몸이 만들어지는 것도 체세포분열을 통해 이루어진다. 예를 들어 불가사리는 체세포분열을 통해 잘린 팔을 재생한다.
- 무성 생식: 일부 생물은 무성 생식 과정을 통해 유전적으로 매우 비슷한 개체를 만든다. 예를 들어 하이드라(hydra)는 싹틈(옛 용어: 출아) 과정으로 무성 생식을 한다. 하이드라의 세포 표면은 체세포분열을 거쳐 싹(bud)이라고 부르는 덩어리를 형성한다. 싹을 가지는 세포에서 체세포분열이 계속 진행되면서 싹은 새로운 개체가 된다. 식물에서도 체세포분열을 통해 영양 번식이라는 무성 생식이 일어난다.
다양성
체세포분열의 형태
진핵생물 세포에서의 체세포분열 과정은 비슷한 패턴으로 진행되지만, 구체적인 과정에서는 3개의 변이가 존재한다. 첫 번째로, 세포 분열 과정에서 핵막이 온전하게 존재하는지 분해되는지에 따라 "닫힌(closed)" 체세포분열과 "열린(open)" 체세포분열로 구별할 수 있고, 핵막이 부분적으로만 분해되는 중간 형태를 "반열린(semiopen)" 체세포분열이라고 한다. 두 번째로, 중기에서 나타나는 방추장치의 대칭성에 따라 구별하기도 한다. 방추장치가 거의 중심에서 축의 방향으로 대칭인 경우를 "정상체세포분열(orthomitosis)"이라고 하고, 방추 장치 모양이 기이하게 좌우대칭인 경우를 "가쪽체세포분열(pleuromitosis)"이라고 한다.세 번째로, 닫힌 가쪽체세포분열의 경우에서 중심 방추의 위치에 따라 구별하기도 한다. 중심 방추가 세포질에 위치하면 "핵바깥"이라고 하고, 중심 방추가 핵에 위치하면 "핵속"이라고 한다.

닫힌
핵속
가쪽체세포분열
닫힌
핵바깥
가쪽체세포분열
닫힌
정상체세포분열
반열린
가쪽체세포분열
반열린
정상체세포분열
열린
정상체세포분열






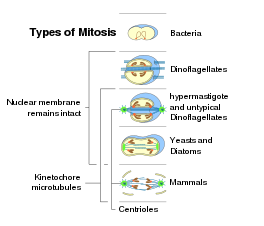
박테리아와 아키아는 핵을 가지지 않기 때문에 핵 분열은 진핵생물 역의 생명체 세포에서만 일어난다. 박테리아와 아키아는 다른 유형으로 분열한다. 예외적으로 모두 닫힌 체세포분열을 하는 섭식구굴착류(Excavata)를 제외하면 각 진핵생물의 상위그룹에서 체세포분열은 닫힌 형태와 열린 형태가 모두 나타난다. 아래는 진핵생물에서 발생하는 체세포분열 형태이다.
- 닫힌 핵속 가쪽체세포분열은 유공충 아문(Foraminifera)와 일부 담녹조 강(Prasinophyceae, 옥시모나드 목(Oxymonad), 아세토스포레아 강(Ascetosporea), 많은 진균들(카이트리디오마이코타 문(Chytridiomycota), 오오마이코타 문(OOmycete), 자이고마이코타 문(Zygomycota), 아스코마이코타 문(Ascomycota)), 일부 방산충 문(Radiolaria)(스푸멜라리아(Spumellaria)와 아칸싸리아(Acantharia))에서 주로 나타난다. 이 유형은 가장 원시적인 유형으로 보인다.
- 닫힌 핵바깥 가쪽체세포분열은 트리코모나디다 목(Trichomonadidia)과 와편모충 문(Dinoflagellata)에서 나타난다.
- 닫힌 정상체세포분열은 규조류(diatom), 섬모충(ciliate), 일부 미포자충(Microsporidia), 단세포 효모와 일부 다세포 진균에서 나타난다.
- 반열린 가쪽체세포분열은 대부분의 정단복합체충 문(Apicomplexa)에서 나타난다.
- 반열린 정상체세포분열은 일부 아메바(Lobosa)와 일부 녹색 편모충(라피도파이트 강(Raphidophyta)이나 볼복스(Volvox))에서 서로 다른 형태로 나타난다.
- 열린 정상체세포분열은 포유류와 다른 후생동물, 그리고 육지 식물, 일부 원생생물에서 나타난다.
오류와 다른 변이들
체세포분열을 하는 동안 오류가 나타날 수 있다. 사람에게서는 특히 초기 배아 발생 과정에서 나타난다. 체세포분열의 각 시기 동안 체세포분열이 정상적으로 일어나게 조절해주는 체크포인트가 존재한다. 그러나 가끔 드물게 실수가 일어날 수 있다. 체세포분열 실수는 염색체 수 하나 이상이 많거나 적은 비정배수체(aneuploid)를 만들 수 있고, 이 경우 암과도 연관이 있다. 초기 사람 배아와 암 세포, 감염되거나 중독된 세포는 3개 이상 딸세포를 만드는 병리학적인 증상을 겪으며, 염색체 구성에 심각한 오류를 야기한다.
비분리는 후기 때 자매 염색분체가 분리되지 않는 현상이다. 하나의 딸세포가 비분리된 염색체에서 2개의 염색분체를 물려받으면, 나머지 세포는 아무 염색체도 물려받지 못한다. 결과적으로 전자의 세포는 그 염색체에 대해 염색체 사본을 3개 가지게 되어 세염색체증(trisomy)이 나타나고, 후자의 세포는 염색체 사본을 1개 가지게 되어 홑염색체증(monosomy)이 나타난다. 가끔 세포가 비분리될 때 이 세포들은 완전한 세포질분열에 실패하여 한 세포에 2개의 핵이 존재하여 이핵 세포(binucleated cell)를 형성하게 된다.
후기 동안 염색체의 이동이 지연되면 후기 지연(anaphase lag)이 일어난다. 이는 방추장치가 염색체에 제대로 부착하지 않았을 때 나타난다. 지연이 일어나는 염색체는 두 딸핵에 들어가지 못하고 소실된다. 그러므로 딸세포 중 하나는 그 염색체에 대해 홑염색체가 된다.
속재복제(endoreduplication)는 염색체가 복제되었지만 세포가 분열하지 않을 때 일어난다. 이는 다배수체(polyploidy) 세포를 만든다. 만약 염색체가 반복해서 복제되면 다섬유 염색체(polytene chromosome)가 만들어진다. 속재복제는 많은 종에서 발생의 정상적인 한 형태로 나타난다. 속체세포분열(endomitosis)은 속재복제의 한 변이로, 세포가 S기 때 염색체를 복제한 뒤 체세포분열에 들어가지만 성숙하지 않은 상태로 체세포분열을 끝내는 과정이다. 2개의 새로운 딸핵을 만드는 대신에, 복제된 염색체는 원래의 핵에 그대로 있는다. 세포는 다시 G1기와 S기로 접어들어 염색체를 또 복제한다. 이 과정은 여러 번에 걸쳐 일어날 수 있어서 복제와 속체세포분열 과정이 끝날 때마다 염색체 수가 늘어난다. 혈소판을 만드는 거대핵세포는 세포 분화 과정에서 속체세포분열 과정을 거친다.
섬모충과 동물의 태반 조직에서 일어나는 무사분열은 부모의 대립유전자를 무작위하게 배분한다.
세포질분열 없이 핵분열만 일어나면 다핵세포를 만든다.
진단 표지자

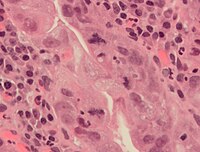
조직병리학에서 체세포분열 속도는 여러 조직 샘플에서 종양의 공격성을 특정하고 진단할 때 사용되는 중요한 지표이다. 예를 들어, 유방암 샘플을 채취하면 일반적으로 체세포분열 횟수를 정량화한다. 체세포분열 횟수는 가장 활발하게 체세포분열을 하고 있는 부위에서 센다. 암은 체세포분열 활성이 매우 높기 때문에 암에서는 이 부위를 확인하기 어렵다. 또한, 비정상적인 체세포분열을 감지하는 것은 진단 표지나 예후 표지로 사용될 수 있다.

암세포에서 나타나는 정상 형태와 비정상 형태의 체세포분열. A, 정상 체세포분열; B, 염색질 다리; C, 다극 체세포분열; D, 고리 체세포분열; E, 분산된 체세포분열; F, 비대칭 체세포분열; G, 지연된 체세포분열; H, 작은 핵. H&E 염색.

관련된 세포 과정
세포 라운딩

동물 조직에서 대부분의 세포는 체세포분열을 할 동안 구 모양이 가깝게 동그랗게 모양이 변한다. 상피와 표피에서 효율적인 라운딩 과정은 방추장치를 적절하게 배열하고 딸세포를 올바르게 위치하는 것과 관련이 있다. 또한, 연구자들은 라운딩이 억제된 세포에서는 방추 결함과 주요 극 쪼개짐에서 결함이 일어나고, 염색체를 효율적으로 잡지 못하는 것을 발견했다. 그러므로 체세포분열 세포 라운딩은 정확하게 체세포분열 하기 위해 필요하다는 것을 알 수 있다.
라운딩을 가능하게 하는 힘은 F-액틴과 마이오신이 수축하는 균질의 세포 겉질로 재배열하는 과정을 통해 발생한다. 그 첫 번째 과정은 세포 주변을 굳게 하는 것이고, 두 번째 과정은 사이기보다 10배 높게 세포 내부의 정수압을 유발하는 것이다. 세포 내부의 높아진 압력은 세포 외부로 힘이 작용하게 하여 주변의 세포나 세포바깥 바탕질을 밀어내면서 세포를 둥글게 만든다. 이 점은 세포끼리 얽매여 있는 조직 시나리오에 특히 중요하다. 압력이 높아지는 것은 포민-매개 F-액틴 핵생성(formin-mediated F-actin nucleation)과 Rho 카이네이스-매개 마이오신 II 수축(Rho kinase-mediated myosin II contraction)에 의존적이다. 두 과정 모두 상류의 신호전달 경로인 RhoA와 ECT2의 지배를 Cdk1의 활성을 거쳐 받는다. 체세포분열에서 라운딩은 그만큼 중요하기 때문에 체세포분열 세포 겉질에서의 분자 성분과 역학은 활발한 연구 분야이다.
체세포 재조합
체세포분열 할 세포를 G1기에 X선으로 피폭시키면, 세포는 두 상동 염색체 사이에서 나타나는 재조합으로 DNA 손상을 복구시킨다. 체세포분열 할 세포를 G2기에 X선으로 피폭시키면, 세포는 두 자매 염색분체 사이에서 나타나는 재조합을 우선적으로 사용해 DNA 손상을 복구시킨다. 재조합에 관여하는 효소의 유전자에 돌연변이가 일어나면, 여러 DNA 손상 물질에 의해 세포가 죽을 수 있는 민감성이 높아진다. 이는 체세포분열 재조합이 잠재적으로 치명적일 수 있는 DNA 손상을 복구시키는 적응 과정을 암시한다.
진화
진핵생물의 체세포분열 과정에 등장하는 액틴이나 튜뷸린 같은 분자들은 원핵생물에도 대응하는 상동 분자가 존재한다. 체세포분열이 진핵생물의 보편적 특징이라는 점에서 체세포분열이 진핵생물 나무의 처음부터 시작되었을 가능성이 크다. 또한 체세포분열은 감수분열보다 덜 복잡하다는 점에서 감수분열이 체세포분열이 등장한 이후에 등장했을 수 있다. 그러나 감수분열이 관여하는 유성 생식은 진핵생물의 원시적인 특징 중 하나인 것은 사실이다. 따라서 감수분열과 체세포분열은 조상 원핵생물의 분열 과정에서 함께 평행하게 진화했을 가능성이 존재한다.
박테리아의 세포 분열에서 DNA가 복제되고 나면, 2개의 원형 염색체는 세포막의 특별한 부위에 부착되어 있다. 반면에 진핵생물의 체세포분열에는 여러 선형 염색체가 존재하고, 염색체의 방추부착점에는 방추의 미세관이 부착한다. 체세포분열의 여러 형태의 관계를 살펴보면 닫힌 핵속 가쪽체세포분열(closed intranuclear pleuromitosis)이 박테리아의 분열과 비슷한 가장 원시적인 유형으로 보인다.
갤러리
체세포분열을 하는 세포를 형광 항체와 염료로 염색시켜서 현미경으로 관찰할 수 있다.

전기 초반: 초록색 가닥으로 보이는 극성 미세관은 온전한 핵 주변에서 바탕질을 형성하고 있다. 파란색은 응축하고 있는 염색체이고, 빨간색은 중심절이다.

전중기 초반: 핵막이 막 사라진 상태이며, 핵막이 사라지면서 미세관은 빠르게 응축한 염색체의 중심절 주변에 위치한 방추부착점과 상호작용할 수 있다.

중기: 중심체는 세포의 양쪽 극으로 이동하여 체세포분열 방추를 형성한다. 염색체는 중기판에 모여 있다.

후기: 방추부착점 미세관은 두 세트의 염색체를 끌어당기며, 극성 미세관은 길어지면서 분열하는 세포를 서로 밀어낸다. 염색체는 최대로 응축된다.

말기: 전기와 전중기 때 일어났던 사건이 거꾸로 일어나며, 세포 주기가 완료된다.





더 보기
참고 문헌
추가 자료
- Morgan, David L. (2007). 《The cell cycle: principles of control》. London: Published by New Science Press in association with Oxford University Press. ISBN 978-0-9539181-2-6.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2002). 〈Mitosis〉. 《Molecular Biology of the Cell》 4판. Garland Science. 2006년 1월 22일에 확인함.
- Campbell N, Reece J (December 2001). 〈The Cell Cycle〉. 《Biology》 6판. San Francisco: Benjamin Cummings/Addison-Wesley. 217–224쪽. ISBN 978-0-8053-6624-2.
- Cooper G (2000). 〈The Events of M Phase〉. 《The Cell: A Molecular Approach》 2판. Sinaeur Associates, Inc. 2006년 1월 22일에 확인함.
- Freeman S (2002). 〈Cell Division〉. 《Biological Science》. Upper Saddle River, NJ: Prentice Hall. 155–174쪽. ISBN 978-0-13-081923-9.
- Lodish H, Berk A, Zipursky L, Matsudaira P, Baltimore D, Darnell J (2000). 〈Overview of the Cell Cycle and Its Control〉. 《Molecular Cell Biology》 4판. W. H. Freeman. 2006년 1월 22일에 확인함.
외부 링크
- A Flash animation comparing Mitosis and Meiosis
- Khan Academy, lecture
- Studying Mitosis in Cultured Mammalian Cells
- General K-12 classroom resources for Mitosis
- The Cell-Cycle Ontology
- WormWeb.org: Interactive Visualization of the C. elegans Cell Lineage – Visualize the entire cell lineage tree and all of the cell divisions of the nematode C. elegans
번역 및 편집: 이동근 (고려대학교 생명과학부 17학번)