

암 줄기세포(Cancer Stem CELL : CSC 또는 Tumor Initiating Cells : TICs)는 (종양이나 혈액암에서 발견되는) 암 세포들로 종양을 생성할 수 있는 능력을 가지는 세포를 말한다. 암 줄기세포는 정상적인 줄기세포와 같은 특징을 갖는데, 구체적으로 특정한 암 샘플에서 발견되는 모든 세포형을 생기게 하는 능력을 지닌다. 즉, 암 줄기세포는 종양을 형성하지 않은 암세포와 다르게 종양형성(tumorigenic)한다. 암 줄기세포는 다양한 세포형에서 줄기세포의 특성인 자기재생과 분화능력을 통해 종양을 발생시킨다. 또한 종양에서 다른집단과 구별되어 새로운 종양을 발생시킴으로써 재발과 전이의 원인이 된다. 그러므로 암 줄기세포를 타겟으로 하여 특이적 치료방법의 발달은 암환자의 생존률의 증가와 삶의 질 개선, 특히 전이성 질병을 가지는 환자들에서 희망이 될 수 있다.
현재 존재하는 암 치료방법은 대부분 동물모델을 기반으로 발달되었으며, 종양을 수축시키는데 효과적이다. 그러나 동물은 사람질병의 완전한 모델을 제공할 수 없는데 실제로, 수명이 2년을 넘지 않은 쥐의 경우, 종양 재발에 대한 연구가 어렵다.
실험 초기단계에서 암 치료방법의 효과는 흔히 종양 덩어리 일부를 절제함으로 측정한다. 암 줄기세포는 종양의 일부이기 때문에 꼭 줄기세포능력에 대한 활성을 갖는 약을 꼭 선택하지 않는다. 이 이론은 전통적인 화학요법(Chemotherapy)이 새로운 세포를 생성하지는 않지만 종양의 대부분을 차지하는 분화되었거나 분화중인 세포를 죽이며, 발생된 암 줄기세포 부분은 그대로 남아 재발의 원인이 된다고 제안한다.
암 줄기세포는 1990년 대 후반 급성 골수성 백혈병에서 존 딕 (John Dick)에 의해 처음 정의 되었다. 2000년대 초반이후로 활발하게 암 연구가 집중되고있다.
종양 증식 모델


종양의 하위 유형에 따라, 종양 개체군 내 세포들은 기능적 이질성 (Heterogenety)을 보인다. 종양은 다양한 증식 및 분화 능력을 가지는 세포로부터 형성된다. 암 세포 중 이러한 기능적 이질성은 종양을 발생시키는 능력의 차이나 이질성을 설명하기 위해 다양한 증식 모델이 제시되었다 : 암 줄기세포 (CSC : Cancer stem cell) 와 클론진화 모델.
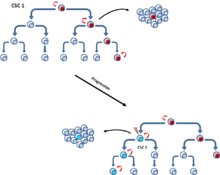
암 줄기세포 모델은 종양 세포의 일부분으로 자기재생 능력을 가지며 다양한 종양 세포 생성이 가능한 종양세포를 나타낸다. 이러한 세포들은 줄기세포와 같은 성질을 갖는 것을 반영하여 암 줄기세포라고 명명된다. 암 줄기세포의 존재와 암 줄기세포 모델은 종양 개체군에서 필연적으로 암 줄기세포가 증식단계 중 최상위에 놓인다고 제시한다. (그림 3) 클론진화모델은 성장에 유리한 조건을 가지는 돌연변이 종양 세포는 일반 다른세포들보다 빠르게 증식한다고 상정한다. 이 우세적 개체군 내 세포들은 암 성장을 발생시키기 위한 비슷한 능력을 가진다 (그림 4).

이러한 두 모델은 상호배타적이지 않은데, 그 이유는 암 줄기세포 자신이 클론 진화를 겪기 때문이다. 그러므로, 만일 하나의 돌연변이가 더 공격적인 특성을 가지게 한다면 이차적으로 우세한 암 줄기세포가 발생할 것이다(그림 5).
기원
암 줄기세포의 기원은 활발하게 연구되고 있는 분야이다. 기원에 대한 대답은 종양 형태와 표현형에 달려있다. 지금까지 암 줄기세포 모델을 사용한다면 종양에서 세포의 기원이 단 하나라는 가설은 증명될 수 없다. 이것은 암 줄기세포가 말기종양에 존재하지 않기 때문이다. 그러므로 심지어 암 줄기세포가 종양을 형성할 수 있는 능력이 있음에도, 세포의 기원으로써 암 줄기세포를 설명하는 것은 부정확한 주장이다.[분류가 필요함]
기원가설들은 줄기세포 또는 간세포 발달 과정 중 돌연변이 생성, 성체줄기세포와 성체간세포에서 돌연변이, 줄기세포와 같은 특성을 지닌 돌연변이를 가진 분화세포를 포함한다. 이러한 이론들은 대부분 종양 세포의 기원에 초점이 맞춰져 있다.
증거
암 줄기세포에 대한 첫 번째 결정적인 증거는 1997년에 발견되었다. 보넷과 딕 (Bonnet 과 Dick)은 표면 표지자 CD38를 발현하지 않고 CD34를 발현하는 백혈구 세포 일부 집단을 분리했다. 저자는 CD34+/CD38- 집단이 비만당뇨(NOD)/중증복합면역결핍병(SCID)쥐에서 종양을 형성하는 능력을 가지는데 이는 조직학적으로 도너와 비슷하다. 고형암에서 암 줄기세포성을 지닌 세포의 첫 번째 증거는 2002년 이후로 나왔다. 사람의 신경교종으로부터 분리되어 클론형성 및 구(Spheroid)를 형성하는 세포를 발견하여 이를 특징지었다. 사람의 피질의 신경교 종양은 생체 외에서 성상교세포와 신경계의 표지자를 발현하는 신경줄기세포와 같은 세포를 포함한다.
암 연구 실험에서, 종양세포를 때때로 종양을 형성하기 위해 실험동물에 주입한다. 시간에 따라 질병이 진행되면 신약의 효험을 알기 위해 테스트되기도 한다. 종양형성은 알려지기 위해 몇 천에서 무수한 많은 세포들이 필요하다. 고전연구에 있어, 이것은 부족한 방법론이나 (즉, 종양세포는 전이되는 동안 그들의 생존력을 잃는다.) 주입된 세포의 특정한 생화학적인 환경인, 미세환경의 중요성에 의해 설명된다. 암 줄기세포 패러다임 지지자는 주입된 세포의 아주 작은 부분인 암 줄기세포가 종양을 형성하는 능력을 가진다고 주장한다. 사람의 급성 골수성 백혈병에서 이러한 세포들의 빈도수는 1000개중 1보다 적다.
추가적인 증거는 조직학으로부터 나왔다. 많은 종양은 이질성을 갖고 숙주의 장기의 고유한 특징을 가지는 다양한 세포형태들을 포함한다. 이질성은 공통적으로 종양 전이성에 의해 갖게되는데 이는 이질성을 갖게하는 세포가 종양 세포의 일반적인 특징인 다양한 세포 형태를 발생시키는 능력을 지녔다는 것이다.
백혈구 줄기세포의 발견은 다른 암에서도 연구를 진행되도록 하였고, 최근에 몇몇 고형암에서 발견되었다.
- 뇌
- 유방
- 대장
- 난소
- 췌장
- 전립선
- 흑색종
- 다발성골수증
- 비흑색종 피부암
수학적 모델

암에 대한 경로를 가정했을 때 세포의 구획을 기반이론으로 하는 등의 수학적 모델은 예측가능 하다. 예를 들어 비정상 세포의 성장은 특이적인 돌연변이 확률을 의미하며,이러한 모델은 성숙세포의 반복된 돌연변이가 비정상적인 자손의 형성하고 암의 생성율을 증가시킨다고 예측할 수 있다. 임상 효과는 아직 확립되지 않았다.
논쟁
암 줄기세포의 존재는 논쟁 속에 있다. 그 이유는 많은 연구들은 암 줄기세포만의 특징을 밝혀내지 못했기 때문이다. 분화된 세포 (헤이플린 한계 "Hayflick Limit"에 의해 제안된)는 무한으로 분열되지 못하지만, 암 세포는 발암에 필요한 많은 돌연변이를 갖고 종양성장을 지속하기 위해 연속적 증식능력과 자기재생 능력이 필수적이다.이는 일반적인 줄기세포의 특징과 비슷하다. 또 다른 논쟁은 암 줄기세포의 기원이다. 일반적인 줄기세포들이 이상조절로부터 생성된 것인지 자기재생능력을 획득한 개체군으로부터 생성된 것인지 논쟁이 되고 있다.
가설
줄기세포 돌연변이
발달과정 중 줄기세포의 특수한 환경에 위치하는 개체군에서의 돌연변이 가설은 발달 중 줄기세포 개체군이 돌연변이가 되고 많은 자손들에게 돌연변이를 유전시키기 위해 재생산된다. 이렇게 생성된 딸세포는 종양이 되기 쉽고 암성 돌연변이가 될 가능성이 증가시킨다.
성체줄기세포
또 다른 이론은 성체줄기세포와 종양생성을 관련시킨다. 이것은 대부분 세포교체율이 높은 조직과 관련이 있다.(피부나 장과 같은). 이러한 조직에서 대부분의 다른 성체줄기세포와 비교하여 성체줄기세포는 그들의 빈번한 세포분열과 긴 수명 때문에 암 줄기세포 기원가설에 후보가 된다. 이러한 결합은 돌연변이가 축적되는 이상의 환경을 생성한다.: 돌연변이 축적은 암 개시단계를 일으키는 일차 인자이다. 이러한 증거들은. 비록 특이적인 암과 특정한 이유와 연결하는 것이 불가능해도 실제적인 현상이 연관성을 보여준다.
역분화
돌연변이 세포의 역분화는 어떠한 세포가 암 줄기세포로 될 수 있을지를 가능성을 제시하면서 줄기세포와 같은 특성을 생성할 수도 있다.
체계
종양 체계의 개념은 종양이 돌연변이 세포의 여러 다른 종류로 이루어진 개체군이라고 주장한다. 이들 모두는 몇몇의 돌연변이를 공유하지만 다양한 특정 표현형 가진다. 하나의 종양은 줄기세포의 여러 가지 형태를 가진다. 어떤 종양은 특정 환경에 대한 최적상태를 가질 수 있고 다른 것들은 덜한 환경에 존재할 것이다. 두 번째의 경우, 치료적 개입 등을 포함한 종양이 그곳에 적응하도록 하면서 다른 환경에서 보다 성공적인 환경일 것이다. 맞다면, 이러한 개념은 암 줄기 세포에 특이적인 치료시스템에 영향을 줄 것이다. 이러한 체계는 암 줄기세포의 기원을 정확히 알아내는데 복잡하고 정교한 시도일 것이다.
암 줄기세포 분리
대부분 사람의 종양에서 암 줄기세포가 확인이 보고되었으며, 연구전반에 정상적인 줄기세포와 비슷한 것을 확인하는 전략으로 많이 이용된다. 이러한 절차에는 형광이용세포분류기/유세포 분석(FACS) 세포 표면의 표지자를 검출하는 항체들과 SP(side population) 분석 (주변 개체군 방법)을 포함한 기능적 접근법이나 Aldefluor 방법을 포함한다. 이러한 암 줄기세포의 많은 결과는 다양한 약들을 투여 되었을 경우 면역결핍 쥐에서 종양발달능력을 평가할 수 있다. 생체 내에서 일어나는 이 방법은 한계희석법이라고 불린다. 낮은 세포 수에서 종양발생을 시작할 수 있는 종양세포의 일부는 추후에 순차적인 종양연구에서 따라 자기재생능력을 실험된다.
암 줄기세포는 또한 다양한 약에 대한 저항성(Multi drug resistance : MDR)을 ABC수송체(ATP-binding cassette)를 통해 회흐스트(hoechst) 염색약의 유출량에 따라 확인될 수 있다. 또 다른 접근방법은 구-형성 (Shpere-forming) 방법이다. 많은 일반적인 줄기세포들은 조혈세포나 조직으로부터의 줄기세포들은 특정한 배양조건에서 3차원 구조의 구를 형성하는데 이는 분화가능하다. 줄기세포와 마찬가지로 암 줄기세포는 뇌나 전립선암으로부터 분리되어 뜬 형태의 구를 형성할 수 있다
이질성 (암 줄기세포 표지자)
암 줄기세포는 다양한 고형암에서 확인되었다. 일반적인 줄기세포에 특이적인 표지자들은 보통 고형암이나 혈액암으로부터 암 줄기세포를 분리하는데 사용된다. 세포 표면의 표지자들은 CD133 (PROM1으로 알려짐), CD44, CD24, EpCAM (상피세포 부착 분자로 상피의 특이적인 항원 : ESA), THY1, ABC수송체B5 (ABCB5),과 CD200을 포함하며, 암 줄기세포가 많은개체군을 분리하는데 유용함이 증명되었다.
CD34 (Prminin 1)은 내피의 전구 세포와 태아의 신경줄기세포에서 CD34+ 줄기세포나 간세포에서 발현되는 5개의 막관통 단백질도메인을 가지는 당단백질이다. 이것은 AC133으로 알려진 당화된 항원결정부를 이용하여 발견할 수 있다
EpCAM (상피세포부착분자인자 :ESA/TROP1)은 대부분 상피 세포의 기저막에서 발현하는 호염성 Ca2+가 풍부한 독립적인 세포 부착 분자이다.
CD90 (THY1)은 원형질막에 연결된 글라이코실포스파티딜이노시톨 당단백질(glycosylphosphatidylinositol glycoprotein)이고 신호전달과정에 관여한다. 이것은 흉선세포와 흉선기질세포 접착에 관여한다.
CD44 (PGP1) 세포의 신호전달에서 이동과 귀소등의 다면발현 역할을 하는 부착분자이다. 이는 히알루론산(hyaluronic acid)에 친밀한 CD44H와 전이 특성을 지닌 CD44V를 포함한 다양한 동형 단백질을 가진다.
CD24(HSA)는 당화된 글라이코실포스파티딜이노시톨-연결부착분자(glycosylphosphatidylinositol-anchored adhesion molecule)로 B와 세포에서 협력적 촉진 작용을 한다.
CD200 (OX-2)는 형태1 막의 당단백질로 T세포와 NK 세포나 대식세포를 포함한 면역세포에 억제신호를 전달한다.
ALDH는 흔한 알데하이드 탈수소효소로 방향족 알데하이드로 산소를 가수분해하여 카복실산(Carboxylic acid) 을 만든다. 예를 들어 레티놀을 생존에 필수적인 레티노산(비타민A)로 전환시키는 역할을 갖는다..
암 줄기세포가 분리되고 확인된 첫 번째 고형 악성종양은 유방암으로 많은 연구가 진행되고 있다. 유방의 암 줄기세포는 CD44+CD24-/LOW,SP ALDH+일부집단에서 많이 발견된다. 유방 암 줄기세포는 특이적인 표현형이 다양하다. 유방암에서 암 줄기세포의 표지자의 발현은 이질성을 띠고, 암 줄기세포집단은 종양 전반에서 다양하다. CD44+CD24- 와 CD44+CD24+ 세포집단은 종양을 만들어내는 세포이다: 그러나 표지자 CD44+CD76f(high)CD133/2(high)를 이용할 때 암 줄기세포가 가장 많이 존재한다.
암 줄기세포는 많은 뇌종양에서 보고되었다. 줄기세포와 비슷한 성질을 갖는 종양세포는 CD133,과 SSEA-1(단계-특이적 배아성 항원-1) EGFR과 CD44을 포함한 세포 표면표지자를 이용하여 확인하였다. 암 종양에서 줄기세포와 비슷한 성질을 갖는 세포를 확인하기 위해 CD133를 이용하는 것은 문제가 있을 수 있다. 그 이유는 종양형성세포는 몇몇 신경교종에서 CD133+와 CD133-에서 발견되고 CD133+일 때 뇌종양이 종양형성능력을 가지지 않는 경우가 존재하기 때문이다.
암 줄기세포는 사람의 대장암에서 보고되었다. 그들은 확인을 위해 CD133.나 CD44, ABCB5,와 같은 세포표면의 표지자와 클론분석과 Aldefluor 방법을 포함한 기능적 분석을 이용했다. 대장암 줄기세포에서 포지티브 표지자인 CD133은 모순되는 결과를 나타냈다. CD133단백질이 아닌 AC133 항원결정부가 대장암 줄기세포에서 특이적으로 발현되고 ,이 발현은 분화과정 중 잃었다. 추가적으로, CD44+ 대장암 세포와 추 CD166를 가진 CD44+EpCAM+ 세포 개체군의 추가적인 부분은 종양접종(생착)의 성공을 증가시킨다.
다양한 암 줄기세포가 전립선 위와 간, 췌장, 신장 및 난소 등 많은 다른 기관 보고되었다.
전립선암에서 종양줄기세포 CD44+a2B1+, TRA-1-60+CD151+CD166+세포는 ALDH+ 개체로써 CD44+세포부분에서 확인되었다. 간의 암 줄기세포로 추정되는 표지자는 CD133+ ALDH+, CD44+ 그리고 태아종양 단백질 5T4+이 보고되었다.
전이성 암 줄기세포
전이는 환자에게 있어 치명적인 종양의 주요 원인이다. 그러나 모든 종양세포가 전이성을 갖지 않는다. 이러한 전이성은 성장, 신생혈관 생성, 침윤 등의 기본적인 과정에 의해 결정되는 인자에 의해 갖게 된다.
상피종양에서 간엽상피전환(epithelial mesenchymal transition : EMT)은 전이에 있어 중요하게 고려된다. EMT와 간엽으로부터 상피형태로 역전환은 배 (Embryo) 발달과정에 포함되어 상피 세포의 항상성 붕괴와 이주성의 간엽의 표현형 습득에 관여한다. EMT는 WNT나 성장인자B를 와 같은 일반적인 경로를 통해 조절된다.
EMT의 중요한 특징은 부착연접부위에서 E-카드헤린(E-cadherin)의 부족이다. 이곳에서 B-카테닌(B-catenin)은 중요한 역할을 한다. 부착연접부위에서 핵으로의 B-카테닌의 위치변화는 E-카드헤린 부족을 가져오며 뒤에 EMT를 유도한다. 핵의 B-카테닌은 E-카드헤린 유전자의 억제자인 SLUG(SNAI2로 알려진)와 같은 EMT-와 관련 있는 타겟으로, 전사적으로 직접 활성화 시킬 수 있다. 더욱이 종양의 미세 환경이나 저산소환경(hypoxia)에서 가지는 기계적(역학적) 성질은 암 줄기세포의 특성과 전이습성에 영향을 끼친다.
EMT과정 중 종양세포는 전이성 암줄기세포의 전구세포일 수 있다.췌장암의 끝에서 침습성 성질을 가지는 CD133+CXCR4+ 세포의(CXCL12 케모카인의 수용체로 SDF1의 리간드로 알려짐)의 일부가 확인되었다. 이러한 세포들은 CD133+CXCR4- 세포보다 두드러진 강력한 전이 활성을 갖는다. 그러나 이러한 세포 둘 다 비슷한 종양발달능력을 지닌다. 더욱이,CXCR4 수용체의 억제는 종향형성능력변화없이 전이능력만 감소시킨다.
암 전이에서 광범위하게 연구된 EMT와 다르게 MET (mesenchymal–epithelial transition)의 역할은 상대적으로 적게 연구되었다. MET는 EMT의 역과정으로, 전이성을 갖는 암성세포가 전이되어 도착된 세포에서 콜로니생성 (colonization) 및 상피세포 가지는 특징과 거리가 있는 다른 기관과 융합되기위한 성질을 얻기위한 안정성과 정착에 관여한다고 믿어진다.
유방암의 CD44+CD24-/낮은 세포는 전이성을 갖는 흉수에서 발견된다. 반대로, 증가된 CD24+세포의 수는 유방암 환자에서 전이성이 거의 없는 곳에서 확인되었다. CD44+CD24-/낮은 세포는 초기에 전이성을 갖고 새로운 부위의 이들은 그들의 표현형을 변화시키고 분화를 제한하는 것이 가능하다. 두단계에서 발현양상의 패턴 가설은 암 줄기세포의 두 가지 형태를 제안한다.-고정적인 암줄기세포(stationary cancer stemcell :SCS)와 이동성을 갖는 암줄기세포(mobile cancer stemcell : MCS). SCS는 종양진행과정 내내 조직에 끼어들어 가거나 분화된 조직내에 계속 남아 있는다.MCS는 종양숙주-인터페이스에 위치한다. 이 세포들은 일시적인 EMT의 획득을 통해 SCS으로부터 얻어진다(그림 7).
암 치료에 대한 영향

암 줄기세포는 질병확인 및 선택적으로 약을 타겟하는 경우, 전이의 예방 및 새로운 치료전략을 포함하여 암 치료에 대한 영향을 끼친다.
정상적인 체세포성 줄기세포(somatic stem cells)는 화학요법에 대한 저항성을 갖는다. 그들은 다양한 펌프(MDR과 같은)를 형성하여 DNA 수선 단백질들과 약을 배출한다. 정상적인 체세포성 줄기세포는 세포교체 속도가 느리다 (화학치료물질은 자연히 빠르게 자기복제하는 세포를 타겟으로 한다) 정상 줄기세포에서 발달된 암 줄기세포는 또한 화학치료에 대한 그들의 저항성을 증가시키는 단백질을 생성할 수 있다 생존하는 암 줄기세포는 재발의 원인으로 종양이 다시 생성될 수 있게 한다. 암 줄기세포는 선택적으로 타겟팅하는 것은 전이와 재발을 막을 뿐만 아니라 종양을 절제하지 않는 공격적인 치료 방법이다. 이 가설은 암 줄기세포 제거함으로써 암이 분화되거나 세포가 죽기 때문에 감소될 수 있다고 말한다. 그러나 종양세포의 어느 부분이 암 줄기세포이고 이를 제거되어야할 필요가 있는지는 명확하지 않다.
연구는 암 줄기세포를 구별할 수 있는 단백질과 유전체측면에서 특이적인 표지자를 찾고 있다. 2009년, 과학자들은 일반적으로 사용되는 화학치료물질인 파클리탁셀(Paclitaxel)보다 100배정도 쥐의 유방암 암 줄기세포를 선택적으로 감소시키는 살리노마이신 (salinomycin)약물을 찾았다. 암 줄기세포의 몇몇 형태는 자가포식(autophagy)을 통해 살리노마이신을 이용한 치료에서 살아남을 수 있다, 또한 특정한 단백질을 감소시키거나 재사용하는 리소솜같은 산성을 띠는 세포소기관을 이용한다. 자가포식억제자 사용은 자가포식을 통해 생존하는 암 줄기세포를 죽일 수 있다.
세포 표면 수용체인 인터루킨-3 수용체-a (CD123)는 급성 골수성 백혈병(acute myeloid leukemia :AML)의 CD34+CD38- 백혈병 줄기세포(Leukemic stem cells: LSCs)에서 과발현하지만 정상적인 CD34+CD38- 골수세포에서는 과발현 되지 않는다. CD123-특이적인 단클론 항체를 AML을 접종시킨 비만당뇨(NOD)/중증복합면역결핍병(SCID) 쥐에 처리한 경우 LSCs가 골수로 돌아가는 것을 손상시키며 이차 수여쥐에서 LSC의 부분을 보함은 전반전인 AML 세포의 골수증식을 감소시킨다.
2015년 연구는 miR-34a와 탄화수소암모늄(NH4HCO3)을 포함한 나노파티클을 패키징해서 쥐의 모델에서 전립선 암 줄기세포에 이들을 전달하였다. 그리고 나서 그들은 적외선 영역 레이저와 가까운 방사능을 처리해주었다. 이것은 나노파티클이 2배에서 그이상으로 크기 증가시켜 엔도솜을 방출시키고 세포내의 RNA을 분산시켰다. miR34a는 CD44의 수준을 감소시킬 수 있다.
경로
암 줄기세포를 타겟하여 새로운 약을 디자인하기 위해서는 세포증식을 조절하는 세포의 기작을 이해하는 것이 필요하다. 이 분야에서 처음 발견은 조혈모세포(Hematopoietic stem cells :HSCs)로부터 만들어졌다. 그들은 백혈병에서 카운터파트너를 변형시켜 생성되는 질병으로 암 줄기세포의 기원이 된다. 많은 기관에서 줄기세포는 백혈병에서 유래한 조혈모세포와 같은 세포경로를 공유한다.
하나의 정상줄기세포는 이를 조절하는 증식 및 분화경로가 비정상적으로 조절되는 경우나 종양단백질 활성을 유도함으로써 암 줄기세포로 변환될 수 있다.
- BMI-1
폴리콤 유전자군의 전사억제자인 BMI-1은 림프종에서 일반적으로 종양유전자(oncogene)를 활성화 시킨다고 알려져 있으며 조혈모세포를 조절한다고 후에 밝혀졌다. BMI-1의 역할은 신경줄기세포에서 묘사된다. 이 경로는 소아 뇌종양의 암 줄기세포에서 활성됨을 보였다.
- Notch
노치(Notch) 경로는 조혈모세포와 신경줄기세포 유방줄기세포를 포함한 여러 가지 세포형에서 줄기세포의 증식을 조절하는 역할을 한다. 이 경로의 구성은 유방암이나 다른 종양에서 종양유전자로 활동한다고 제안되고 있다.
노치 신호전달경로에서 특정한 부분은 Hes3 전사인자를 포함하는데 이는 교모세포종로부터 얻어진 암 줄기세포 특성을 지닌 많은 수의 배양된 세포를 조절한다.
- 소닉 헤지호그(Sonic hedgehog) 와 WNT
이 발달된 경로는 줄기세포 조절자이다. 소닉 헤지호크와 WNT 경로는 일반적으로 종양에서 과하게 활성화 된다. 이것은 종양성장유지에 필수적이다. 그러나 소닉 헤지호그에 의해 조절되는 Gli 전자인자는 이름을 따온 신경교종에서 높게 발현된다. 이 두 개의 경로에는 어느정도 겹치는 부분(간섭)이 존재하여 보통 같이 활성화 된다.이것은 보편적이다. 예를 들어 대장암의 경우 헤지호그 신호전달이 wnt 경로를 막는것처럼 보인다.
소닉 헤지호그 차단제인 사이클로파민(Cyclopamine)를 이용할 수 있다. 수용성인 사이클로파민은 암 치료에 더욱 효과적일 수 있다. DMAPT는 AML(백혈병)과 골수종 전립선암에 위한 파테놀리드(Parthenolide :산화적 스트레스를 유도하고 NF-kB 신호전달을 억제하는의 수용성 파생물이다.
텔로머레이스(telomerase)는 암 줄기세포 생리에서 연구과제이다. GRN163L(Imetelstat : 텔로머레이스 억제제)는 최근에 골수줄기세포를 타겟으로 하는 시도가 최근에 시작되었다.
WNT 신호전달은 정상 수용체가 신호를 받지 않았음에도 불구하고 영구적으로 활성을 지니게 되는 후속의 종양유전자나 종양억제 유전자에 돌연변이 생기는 경우, 지속적인 자극이 필요하지 않을 것이다. B-카테닌은 단백질 TCF4와 같은 전사인자와 결합하여 필수적인 유전자들을 활성화 시킨다. LF3는 강하게 세포주의 생체 외에서 이 결합을 억제하여 건강한 세포에 영향을 주지않고, 쥐 모델에서 종양성장을 감소시킨다. 이것은 복제를 막고 이동능력을 감소시킨다. 치료 후 암 줄기세포가 아닌 세포가 남게된다. 이러한 발견은 알파스크린(AlphaScreen)과 효소 결합 면역 침강 분석법(Enzyme-linked immunosorbent assay :ELISA)기술을 포함하는‘합리적 의약 설계’의 생산물이다.
암줄기세포를 이용한 3D 모델의 개발
구형태로 자란 암 줄기세포의 단층막은 MDA-MB 231세포보다 더 나은 성장률을 보여준다. 이것은 암줄기세포가 3D의 스페로이드 형태의 효과성을 보여준다. CD44는 MDA-MB231의 2D 배양과 비교하여 스페로이드에서 발현이 증가되었다. ALDH1은 유방암의 줄기세포의 중요표지자로 3D로 키운 유방암 줄기세포와 MDA-MB231에서 많이 발현되었다. 반면에 2D로 자란 MDA-MB 231 세포와 암줄기세포에서는 덜 발현되었다.
스페로이드 형태로 자란 암 줄기세포는 성장률이 좋으며 이는 암 줄기세포 배양할 때 3D 스페로이드 형태로 키우는 것이 효과적임을 보여준다. BCSC 출현율과 임상결과를 보면 유방암 세포의 전이와 약에 저항성이 암 줄기세포가 중요한 역할을 한다.
참고 문헌
인용
- Polyak K, Weinberg RA (April 2009). “Transitions between epithelial and mesenchymal states: acquisition of malignant and stem cell traits”. 《Nature Reviews Cancer》 9 (4): 265–73. doi:10.1038/nrc2620. PMID 19262571.
- Sánchez-García I, Vicente-Dueñas C, Cobaleda C (December 2007). “The theoretical basis of cancer-stem-cell-based therapeutics of cancer: can it be put into practice?”. 《BioEssays》 29 (12): 1269–80. doi:10.1002/bies.20679. PMID 18022789.
- Gao JX (2008). “Cancer stem cells: the lessons from pre-cancerous stem cells”. 《Journal of Cellular and Molecular Medicine》 12 (1): 67–96. doi:10.1111/j.1582-4934.2007.00170.x. PMID 18053092.
- Yanyan Lia, Tao Zhang (May 2014). “Targeting cancer stem cells by curcumin and clinical applications”. 《Cancer Letters》 346 (2): 197–205. doi:10.1016/j.canlet.2014.01.012.
외부 링크
- European Cancer Stem Cell Research Institute A new Institute dedicated to research into cancer stem cells and related work.
- Cancer Stem Cell News A blog of news items related to cancer stem cells, with an emphasis on recent research and articles that are openly accessible
- Exploring the role of cancer stem cells in radioresistance Abstract of a review by Michael Baumann, Mechthild Krause, Richard Hill, "Nature Reviews Cancer " 2008(Jul); 8(7) 545-54
- "A Tumor's Lifeblood", Jessica Gorman, CR magazine, Summer 2006
- "Cancer Stem Cell Scientific Literature Review", UMDNJ Stem Cell Research and Regenerative Medicine, June 17, 2006
- "Stem cells may cause some forms of bone cancer", News-Medical.Net, December 7, 2005
- "The Bad Seed: Rare stem cells appear to drive cancers" Archived 2008년 4월 18일 - 웨이백 머신, Science News Online, March 20, 2004
- "The Real Problem in Breast Tumors: Cancer Stem Cells", Genome News Network, March 7, 2003
- Differentiation Therapy - A Different Approach to Treating Tumors (from Beaker Blog)
- Characteristics of Cancer Cells Cancer Inform Blog
- Cancer stem cells may be cause of brain tumors (research of John A. Boockvar)