
| 소뇌 | |
|---|---|
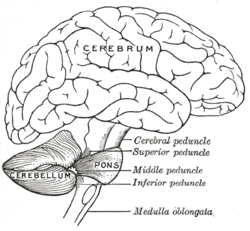 소뇌와 교뇌가 강조되어있는 뇌 도면
| |
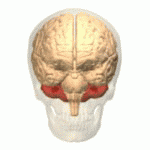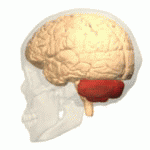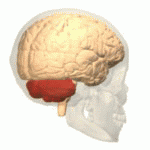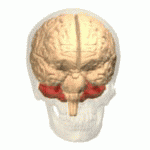 소뇌가 붉은색으로 표시되어 있다
| |
| 정보 | |
| 상위 구조 | 후뇌 |
| 식별자 | |
| 라틴어 | Cerebellum |
| MeSH | D002531 |
| NeuroNames | 643 |
| NeuroLex ID | birnlex_1489 |
| TA98 | A14.1.07.001 |
| TA2 | 5788 |
| FMA | 67944 |
소뇌(小腦, cerebellum)는 척추동물 대부분의 후뇌에서 도드라게 관찰되는 뇌의 일부분이다. 대개 대뇌보다는 작지만, 코끼리고기과 동물과 같은 일부 동물들은 대뇌보다 소뇌가 더 큰 경우도 있다. 감각 인지의 통합과 근육의 조정과 제어에서 중요한 역할을 담당하는 것으로 알려져있다. 그 이외에도 주의, 언어와 같은 인지 기능과 두려움 조절, 쾌락 반응 등에 관련되어 있는 것이 알려져 있다. 운동기능의 경우 운동 신호를 직접 내려주는 중추가 아닌, 척추를 통해 들어온 감각 정보와 기타 뇌 영역의 신호들을 종합하여 정밀한 운동기능을 수행하거나 협응하여 수행한다.
해부학적으로는 대뇌 하부에 분리되어 존재한다. 불규칙적으로 분포한 대뇌 피질의 고랑들과는 다르게, 소뇌 피질의 고랑들은 서로 평행하여 규칙적으로 배열되어있다. 소뇌 피질에는 여러 종류의 신경 세포가 규칙적으로 배열되어 있는데 그 중 푸르키녜 세포와 과립세포가 특히 중요한 기능을 한다. 이들이 수행한 작업들은 대부분이 소뇌 중심부 백질에 위치한 소뇌심부핵을 통과해 다른 영역으로 전달된다.
구조
소뇌는 교(橋) 등쪽의 제4뇌실에 들씌워지듯이 존재하는 큰 구조로, 인간의 경우 가로 10cm, 세로 5cm, 높이 3cm, 무게는 약 150g 정도이다.
소뇌의 위쪽은 대뇌 반구의 후두엽에 접해 있지만 거기에는 경막이 들어 있어 두 부위를 구분한다. 이 막은 지붕처럼 소뇌를 덮고 있기 때문에 '소뇌 텐트'라고도 한다. 소뇌는 양쪽으로 크게 부풀려 있는데, 이 부분을 '소뇌 반구'라고 하며, 좌우 소뇌 반구에 낀 가느다란 중앙부를 '충부(蟲部)'라고 한다. 충부와 소뇌 반구는 형태적으로는 연속해 있으나 그 기능은 완전히 다르다. 또 진화적인 관점으로 보면 충부는 역사가 긴 부분으로 '고(古)소뇌'라고도 한다.
소뇌는 매우 단단하게 접혀진 구조로 되어 있으며 피질은 쭈글쭈글한 주름(소뇌회)으로 이루어져 있는데, 대뇌 반구와는 다르게 거의 평행으로 가로주름 모양으로 있다. 주름 사이의 홈은 얕은 것이 주로 있지만 곳곳에 아주 깊이 패어있고, 내부에서 나뭇가지 모양으로 갈라져있다. 소뇌 백질의 깊숙한 안쪽으로 겹겹이 쌓여져있는 핵들이 존재하며 가장 아래 부분은 액체로 가득찬 뇌실이 존재한다.
소뇌 반구는 고등동물에 발달한 부분으로, 이를 '신(新)소뇌'라고 부르기도 한다. 어류나 조류에도 소뇌는 발달해 있고 크기도 크지만 이들은 모두 고소뇌만으로 되어 있다. 소뇌와 기타 뇌와는 상·중·하의 세 가지 소뇌각(脚)에 의해 연락을 하는데, 이들은 모두 신경 섬유 다발이다. 이 가운데 상소뇌각은 소뇌핵에서 나와 중뇌·간뇌 등으로 가는 섬유를 주로 함유하는데, 중소뇌각은 대뇌 피질에서, 하소뇌각은 연수·척수에서 소뇌 피질에 들어오는 섬유를 주로 함유하고 있다.
내부 구조
소뇌의 내부 구조는 가장 바깥층에 신경 세포 집단인 '피질'(소뇌 피질), 그리고 여기서 나오는 신경 섬유 다발로 된 '수질'이 있으며, 가장 내부에는 신경 세포 덩어리인 '소뇌핵'이 있다.
소뇌 피질
두개의 반구로 이루어진 소뇌 피질은 모든 부분이 균등한 구조를 하고 있으며 중앙부인 충부(Vermis)를 넘어 양 옆에 있는 반구로 연장된다. 피질의 표면이 매우 밀접하게 접혀 소뇌회(folia)를 이루고있다. 대뇌 피질과는 매우 다른 모양으로 소뇌 피질은 뚜렷하게 3층으로 나누어 진다.
과립층(Granular layer)
과립층은 소뇌 피질의 가장 안쪽 부분으로 이끼섬유를 통해 외부로부터 입력신호가 들어오는 부분이다. 과립층의 가장 큰 특징은 지름 5~8um 정도의 작은 과립 세포들이 밀집되어 있는 것이다. 과립 세포는 핵막 주위에 염색과립을 가지고 있어서 소뇌 피질을 염색하면 매우 진하게 염색이 된다. 이 과립세포들의 수는 매우 많아 인간의 경우 중추신경계의 신경세포들 중 50% 이상을 차지한다. 과립세포의 축삭들은 과립층에서 수직으로 상승하여 분자층까지 들어간 후 여기서 2갈래로 갈라져서 소뇌 피질의 평행방향으로 뻗어 평행섬유를 이룬다. 이 외에도 골지 세포, 골홑극붓털 세포, 루가로 세포들이 존재하여 신경망을 형성한다. 구체적으로는 이끼섬유들이 과립 세포, 골홑극붓털 세포, 골지 세포에 글루타민산이 전달물질인 시냅스를 형성하며, 골홑극붓털 세포는 다시 과립세포에 다시 글루타민산 시냅스j를 형성한다. 골지 세포 또한 역시 과립 세포에 감마 아미노뷰티르산(GABA)을 신호물질로 하는 시냅스를 형성한다. 루가로 세포들은 푸르키네 세포들로부터 GABA 입력을 받고 다시 골지 세포들에게 축삭을 보내 GABA 시냅스를 형성한다.
푸르키네 세포층(Purkinje cell layer)
푸르키네 세포층은 과립층 바로 바깥쪽에 위치한다. 여기에는 소뇌 피질에서 눈에 띄는 '푸르키네 세포'의 세포체들이 자리한다. 지름 15-20um의 세포체가 푸르키네 세포층에 늘어서 있고, 여기서 나오는 축삭(원심성)은 피질에서 나와 소뇌핵으로 향하는데, 이것이 소뇌 피질에서 소뇌핵으로의 유일한 신경섬유이다. 푸르키네 세포의 수상 돌기는 가늘게 가지가 갈라지면서 부채꼴로 퍼져 분자층으로 뻗어나간다. 여기서 올리브핵에서 뻗어나온 등정섬유들이 각 푸르키네 세포마다 하나씩 붙어 글루타민산 시냅스를 형성하고 수상 돌기를 따라 계속 시냅스를 형성해가며 분자층으로 올라간다. 또한 분자층의 바구니세포로부터 뻗은 축삭이 푸르키네 세포 축삭의 첫번째 마디와 세포체를 바구니처럼 감싸면서 GABA 시냅스를 형성한다.
분자층(Molecular layer)
분자층은 소뇌 피질 가장 바깥에 위치한다. 이 부분은 과립층의 과립 세포들의 축삭인 평행섬유로 두텁게 뒤덮여있다. 여기서 과립세포의 축삭들은 푸르키네 세포와 골지 세포의 가지돌기들에 글루타민산 시냅스를 형성한다. 또한 이 층에는 바구니 세포(basket cell)와 성상세포(stellate cell)가 존재하고. 이 세포들의 가지돌기는 푸르키네 세포와 같이 소뇌피질에 수직으로 뻗어있어 과립세포의 축삭들과 글루타민산 시냅스를 형성한다. 바구니 세포와 성상 세포는 각각 푸르키네 세포의 축삭 및 세포체와 푸르키네 세포의 가지돌기에 GABA 시냅스를 형성한다. 하나의 바구니세포는 소뇌피질의 횡단면에서 10개 정도의 푸르키네 세포와 신경접합을 이룰 수 있다. 그 외에도 성상세포들은 자기들끼리 GABA 시냅스를 형성한다.
세포 구성
푸르키네 세포(Purkinje cell);조롱박 세포
분자층과 과립층 사이에 두 층의 세포층을 따라서 늘어서 있다. 그 가지돌기는 보통 세포체의 말초부에서 1~2개의 1차 가지돌기로 시작하여 피질의 표면을 향해 광범위하게 퍼져 그 끝이 연막 바로 아래까지 위치한다. 푸르키네 세포의 폭은 약 30μm이고 직경은 약 70μm이다. 가지돌기 가시(spine)가 약 180000개가 존재하여 가지돌기 수용면이 매우 넓어지며 사람에게 푸르키네 세포는 약 15X106개로 추정된다. 푸르키네 세포의 축삭, 특히 충부에서 나오는 것은 전정신경외측핵에 도달하는 것이 있다. 푸르키네 세포의 축삭은 측지를 내서 골지세포나 바구니세포의 가지돌기와 시냅스결합을 한다.
골지세포 (Golgi cell)
골지세포는 과립층의 표층 가까이에 있고 긴 방사상의 가지돌기를 모든 방향으로 펼치고 있다. 이들 가지돌기의 대부분이 분자층으로 들어가지만 과립층에만 존재하는 것도 간혹 있다. 골지 세포의 축삭은 세포체의 아래부분에서 시작하는데 이때 세포체에서 나오자마자 미세하게 분지하여 빽빽한 섬유속을 이루어 과립틍에 분포한다. 골지세포의 축삭은 이끼섬유와 함께 과립세포의 가지돌기에 시냅스 결합하여 이른바 소뇌사구체를 형성한다. 골지세포로 들어가는 흥분충동은 주로 분자층의 평행섬유와 푸르키네 세포의 축삭측지에 의해 전달된다.
과립세포 (Granule cell)와 이끼 섬유(Mossy fiber)
과립세포는 중추신경계 중에서 가장 작은 신경세포이며 소뇌피질의 최심층 즉 과립틍에는 무수한 과립세포가 밀집해 있다. 과립세포의 직경은 5~7μm이고 핵은 원형으로 염색이 잘되고 얇은 세포질층에 둘러싸여 있다. 과립세포는 각각 4~7개의 가지돌기를 방사사으로 펼치고 있다. 가지돌기의 갈구리 모양의 종말부는 소뇌 사궁체에 들어가 여기서 이끼섬유와 시냅스 연결한다. 이끼 섬유는 각 20~30개의 사구체에 가지를 보내며 사구체는 각각 약 15개의 다른 과립세포로부터 가지돌기를 받고 있다. 따라서 각 1개의 이끼섬유가 적어도 300개의 과립세포와 연결된다. 과립세포의 축삭은 분자층에 수직으로 상행하여 여기서 2개의 가지로 갈라진다. 이렇게 갈라진 가지는 평행섬유(parallel fiber)로 되어 소뇌회의 장축에 평행해서 2~3mm 뻗어있다. 과립층의 심부에 있는 과립세포의 축삭은 분자층의 심부에서 갈라져 푸르키네 세포의 가지돌기의 가까운 부위의 가지돌기극과 시냅스 결합한다. 분자층에 있어서 평행섬유는 마치 전화선의 굵은 다발 같이 푸르키네 세포의 가지돌기에 수직으로 걸려 있다. 평행섬유는 약 220개의 푸르키네 세포를 통과하고 각 푸르키네 세포는 3X105~4X105개의 평행섬유에 의해 관통된다.
바구니 세포(Basket cell)과 성상세포(Stellate cell)
바구니세포와 성상세포는 분자층에 있으며 이들 세포의 가지돌기는 푸르키네 세포의 가지돌기와 마찬가지로 시상면에 분포한다. 축삭은 세포체의 아래에서 시작하여 시상면으로 푸르키네 세포의 가지돌기 분지와 평행하게 갈라져 위치한다. 바구니 세포의 축삭은 세포체에서 약간 떨어진 곳에서 하행지를 내고 이들은 많은 시냅스로 푸르키네 세포에 종지하지만 이보다 먼저 푸르키네 세포 축삭을 바구니 모양으로 둘러싼다. 1개의 바구니 세포의 축삭은 시상방향으로 달려서 약 12개의 푸르키네 세포와 결합하고 이것과 수직방향으로 2~3개의 푸르키네 세포와 결합한다. 그러므로 바구니세포는 분자층의 심부에 있는 특수화 된 성상세포로 볼 수 있다.
오름섬유(Climbing fiber)
오름섬유는 뇌줄기(brain steam)와 반대편 아래올리브핵(inferior olivary nucleus)을 통하여 푸르키네 세포로 신호를 전달한다. 오름섬유는 소뇌피질로 들어가기전 약 10개의 가지로 나누어져 푸르키네 세포의 가지돌기와 수 백개의 시냅스를 형성한다. 팽행섬유가 푸르키네 세포로 많은 신호를 보내는 반면, 오름섬유는 푸즈키네 세포로 하나의 신호만 보낸다. 오름섬유의 활동전위는 푸즈키네 세포의 활동전위를 유도한다.
역할 및 작용
소뇌의 들신경(afferent fiber)은 날신경(efferent fiber)에 비해 약 3배 정도로 많다.
구심성 회로(Afferent pathway)
소뇌로 들어오는 섬유는 아래소뇌다리(Inferior cerebellar peduncle), 중간소뇌다리(Middle cerebellar peduncle), 위소뇌다리(Superior cerebellar peduncle)이 있으며, 주로 아래소뇌다리와 중간소뇌다리를 통한다.
다리뇌소뇌로(Corticopontocerebellar pathway)
피질교뇌 섬유(Corticopontine fiber)는 대뇌 피질의 이마엽, 두정엽, 측두엽, 뒤통수엽의 신경세포가 방사관(Corona radiata)과 내포(Internal capsule)를 지나서 교핵, 다리뇌핵(Pontine nuclei)에 도착한다.
올리브소뇌로(crebro-oliocerebellar pathway)
피질올리브 섬유(cortico-olivary fiber)는 대뇌 피질의 이마엽, 두정엽, 측두엽, 뒤통수엽의 신경세포가 방사관과 내포를 통과하고 소뇌 양 쪽에 존재하는 올리브핵에 도착한다.
그물소뇌로(cerebroreticulocerebellar pathway)
대뇌 피질 중 감각 영역에 존재하는 신경세포, corticoreticular 섬유가 연수의 정중곁그물핵(Paramedian reticular nucleus)과 가쪽그물핵(Lateral reticular nucleus)으로부터 소뇌의 양쪽에 존재하는 아래소뇌다리 핵 (Inferior olivary nuclei)로 들어온다.
원심성 회로
소뇌 피질에서 나가는 모든 원심성 회로는 푸르키네세포의 축삭을 통해서 나간다. 소뇌 피질에서 나가는 원심성 섬유는 적색 핵(red nucleus), 시상(thalamus), 전정 복합체(vestibular complex), 망양체(reticular formation)와 연결되어 있다.
둥근쐐기적색로 (Globose-Emboliform-rubal pathway)
구성핵(둥근핵)과 전상핵(쐐기핵 or 마개핵)에서 나오는 축삭은 위소뇌각을 지난다. 그 후 적색핵에서 연접 결합을 한다.
치아시상로 (Dentothalamic pathway)
치상핵에서 나오는 원심성섬유는 상소뇌각을 통해 소뇌를 지나 적핵의 바로 옆을 상행해서 시상에서 연접하고 일부는 적핵에 연접 결합한다. 상행섬유는 대부분에서 하행성 측지가 기시해 뇌간 피개부를 하행에서 피개부의 망상체, 특히 정중방망상핵과 연접 결합한다. 상행섬유군의 대부분은 시상 외측 또는 전복측핵과 연접 결합하고 대뇌피질 제 4, 6영역으로 투사된다.
중위핵→반대측 적핵 대세포부→교차성 적핵 연수로, 적핵척수로 외측핵→반대측 적핵→비교차성 적핵하올리브핵
꼭지안뜰로 (Fastigial Vestibular pathway)
실정핵(꼭지핵)의 저부에서 나오는 섬유군은 정중선을 교차한 후 상소뇌 각 주위를 돌아 하행성 섬유는 반대측의 전정신경핵이나 뇌간망상체(그물체)에 연접 결합을 한다.
꼭지그물로 (Fastigial Reticular pathway)
실정핵의(꼭지핵) 저부에서 나오는 섬유군은 정중선을 교차한 후 상소뇌 각 주위를 돌아 상행 섬유는 시상에 도달하여 연접 결합을 한다.
기능
소뇌의 기능은 행동장애를 검사하는 과정에서 밝혀졌다. 소뇌가 손상될 경우 미세한 움직임 조절에 문제가 발생한다. 소뇌의 손상 여부를 알아보는 검사 중에는 팔을 양 옆으로 뻗은 후 양쪽 손가락이 만나는데 걸리는 시간을 측정하는 방법이 있는데, 소뇌가 손상된 사람의 경우 손가락이 만나는 시간이 오래 걸리지만 소뇌가 건강한 사람의 경우 빠르게 손가락 끝이 닿는다. 1990년까지 소뇌의 기능이 운동조절으로만 알려졌지만 이후 언어, 주의력 등의 새로운 기능이 밝혀졌다. 뇌 부위간의 관계연구에서 대뇌피질의 비운동부위와 소뇌가 상호작용을 발견했으며, 운동능력과 관련이 없는 환자들에서 소뇌의 손상이 확인되었다. Kenji doya는 "소뇌가 많은 수의 독립적인 구조들로 구성되어있고 그 구조들은 기하학적이고 규칙적으로 이루어져있기 때문에 동일한 연산작용을 수행한다. 그래서 소뇌는 감정, 행동과 같은 서로 다른 특성을 조절할 수 있다"라 주장했다.
운동조절
소뇌의 작용은 골격근의 활동 조절을 하는 것이다. 어떤 운동을 할지, 그러기 위해서는 몇 가지 근육을 어떻게 사용해야 하는지에 대한 계획은 대뇌 피질의 전두엽(6영역)에서 세워지는데, 실제로 운동이 시작되고 나서 그 계획대로 여러 활동이 실현되도록 피드백 기구로 관여하는 것은 소뇌이다. 그래서 소뇌는 '추체 외로'라는 운동 명령 전달 경로에 관계하고 있다.
이 조절 양식은 두 가지로 크게 나눌 수 있다.
- 첫째는 주로 내이의 평형 감각 기관에서 오는 정보를 받아들여 몸의 중력과의 관계나 운동에 의한 가속도 변화 등에 대처하여 적절한 반사 운동을 하도록 조절하는 것이다. 평형 감각 기관에서 나온 전정 신경은 일부는 직접적으로, 일부는 교(橋) 등쪽에서 다른 신경과 결합한 뒤 소뇌 피질의 충부에 도달한다. 이 부분이 상처를 입으면 현기증이 나거나 몸의 평형이 깨진다. 또 이 부분에서의 명령은 교의 등쪽이나 연수의 신경핵 또는 망양체에 보내진 뒤 곧바로 척수로 하행하기 때문에 대뇌 피질 등이 관여할 여지는 거의 없으며, 조절은 완전히 무의식적으로 행해진다.
- 둘째는 골격근에 의한 수의 운동이 실행될 때 근육의 수축 정도나 각종 근육의 협력을 위한 조절을 하는 것이다. 이것은 다시 둘로 나뉜다. 하나는 근방추(筋紡錘)·건(腱)방추 또는 근육 자체에서 그 활동 상태 정보를 받기 때문에 척수나 연수 등을 거쳐 소뇌 반구(신소뇌)의 전반부에 도달한다. 또 하나는 대뇌 피질·시상(간뇌) 등에서 운동의 목적이나 계획 등의 정보를 받아들이는 것으로, 교핵을 통해 소뇌 반구 후반부에 도달한다. 이 부분들이 손상을 입으면 걸을 때 비틀거리거나 손을 목적물에 곧바로 뻗을 수 없거나 한다. 이것을 '소뇌성 운동 실조'라고 한다.
소뇌 피질에서 내보내는 명령은 소뇌핵을 통해 주로 간뇌나 중뇌로 보내져 거기에서 추체 외로에 연락한다. 따라서 제1경로보다 복잡하며, 그만큼 다른 방면에서 정보가 개입할 여지를 남기고 있다. 또 소뇌핵에서는 대뇌 피질에도 간접적이나마 연락이 있으며, 조절 기구의 일부는 의식할 수 있다는 것을 나타낸다.
학습
소뇌는 운동학습에 중요한 역할을 하며, 행동을 개시하기 전 학습된 미세 움직임을 조절하는 역할을 한다. 하지만 미세조절을 하기 위한 운동학습을 소뇌가 직접 관여를 하는 것인지, 단지 뇌의 다른 부위에 운동학습에 대한 신호를 전달하는 것인지에 관한 여부는 정확히 밝혀지지 않았다.
병리학
- 소뇌충부증후군 : 소뇌충부의 수질아세포종을 원인으로 한 증상이다. 편엽소절엽이 침범된으로써 전정계와 관련된 증상과 징후들이 나타난다. 소뇌충부는 하나뿐으로 중앙선상의 구조물들에 영향을 주기 때문에 사지를 제외한 머리나 체간에서 근육장애가 나타난다. 앞이나 뒤로 쓰러지는 경향이 있으며 머리를 지탱하거나 똑바로 서있기가 힘들다. 또한 몸통을 똑바로 세우는 것도 힘들다.
- 언어장애 : 구음장애는 소뇌질환에서 푸두근육의 운동실조로 인해 발생한다. 말을 끊는 것이 경련적이며 음절들은 대개 서로 분리되어 말하게 된다. 말이 폭발하듯 터지는 경향이 있으며 음절드이 중복되는 수도 있다. 중요한 것은 소뇌질환에서는 마비나 감각변화가 없다는 점이다.
- 안구운동장애 : 안구진탕은 안구근육의 운동실조로서 안구가 주기적으로 진동하는 현상이다. 안구가 수평방향으로 편위되었을 때 더 쉽게 나타난다.
- 전엽증후군 : 체위반사장애와 제뇌경직이 나타나고 이들의 경우 사람에서는 전엽증후군은 나타나지 않고 같은 부위의 장애가 심한 만성알콜중독에 의한 소뇌변성예나 만발설 소뇌피질위축증에서 다리에 심한 실조즈앙이 나타난다. 어느것이든 소뇌증상의 국재는 반드시 명확하지 않다.
- 신소뇌증후군 : 신소뇌의 장애에 의해 같은 쪽의 근육긴장저하, 공동운동불능 및 변환운동장애 들이 나타난다. 치상핵까지 장애되면 의도진전이 더해진다.
역사
소뇌는 겉모양에 독특한 특징을 지니고 있기 때문에 최초의 해부학자들도 이에 대한 기록을 따로 남겼다. 아리스토텔레스와 갈레노스에 의해 인용된 헤로필로스는 소뇌를 뇌(고대 그리스어: ἐγκέφαλος)와 반대된다는 뜻으로 파렌케팔리스(고대 그리스어: παρεγκεφαλίς)라 불렀다. 갈레노스는 소뇌를 운동신경의 중추로 보았다.
이후 특별히 탐구되지 않다가 르네상스 시대에 다시 주목된다. 안드레아스 베살리우스가 소뇌에 대해 가벼운 논문을 남긴 뒤 1664년 토마스 윌리스가 철저하게 서술되었다. 해부학적 관찰을 넘어 기능적 발견이 시작된 것은 19세기 초의 일이다. 1809년 루이지 롤란도는 소뇌가 손상되면 운동장애가 발생한다는 사실을 발견했다. 마찬가지로 19세기 초반 장피에르 플로랑은 동물 연구를 통해 소뇌가 손상되어도 움직일 수는 있지만 운동에 대한 조정능력을 상실한다는 것을 발견했고, 병변이 크지 않다면 기능을 일부 회복하는 것도 가능함을 밝혀냈다. 20세기가 되면서 소뇌의 주기능이 운동조절과 관련 있다는 것이 일반화되었고 20세기 전기에 들어 인간의 소뇌질환과 관련된 몇몇 임상적 증상들도 자세히 소개되기 시작했다.
같이 보기
참고 문헌
- Clarke E, O'Malley CD (1996). "Ch. 11: Cerebellum". The Human Brain and Spinal Cord (2nd ed.). Norman Publishing. p. 629. ISBN 0-930405-25-0.
- Ito M (2002). "Historical review of the significance of the cerebellum and the role of Purkinje cells in motor learning". Ann. N. Y. Acad. Sci. 978: 273–288. Bibcode:2002NYASA.978..273I. doi:10.1111/j.1749-6632.2002.tb07574.x. PMID 12582060.
- Fine EJ, Ionita CC, Lohr L (2002). "The history of the development of the cerebellar examination". Semin. Neurol. 22 (4): 375–84. doi:10.1055/s-2002-36759. PMID 12539058.
- 신경해부학 제 5판, 김성수, 박승택, 서영석, 안의태, 원무호, 이인환, 이화동, 허영범, 한후재, 황세진, 정문각(1996)
- 신경해부학 제 27판, 허영범 외 공역, 범문에듀케이션(2013)