
단백질의 1차 구조(Protein Primary Structure)는 펩타이드 또는 단백질에서 아미노산의 선형 서열이다. 통상적으로 단백질의 1차 구조는 N 말단에서 C 말단으로 생성된다. 단백질 생합성은 세포의 리보솜에 의해 이루어진다. 실험실에서도 펩타이드를 합성할 수 있다. 단백질 1차 구조는 직접 서열을 분석하거나 DNA 서열로부터 추론된다.
형성
생물학적
아미노산은 펩타이드 결합을 통해 중합되어 긴 골격을 형성하며, 다른 아미노산 곁사슬이 돌출되어있다. 생물학적 관점에서, 단백질은 세포의 리보솜의 번역에 의해 생성된다. 또한 일부 유기체는 비리보솜 펩타이드 합성에 의해 짧은 펩타이드를 만들 수 있는데, 이는 20개의 표준 아미노산 이외의 아미노산을 사용하며 고리화, 변형, 가교될 수있다.
화학적
펩타이드는 다양한 실험실 방법을 통해 화학적으로 합성될 수 있다. 화학적 방법은 전형적으로 생물학적 단백질 합성과 반대 순서로 펩타이드를 합성한다(C 말단에서 시작).
표기법
단백질 서열은 일련의 문자로 표시되며, N 말단에서 시작하여 C 말단까지 아미노산이 열거된다. 세 글자 약어(Three Letter Codes) 또는 한 글자 약어(One Letter Codes)를 사용하여 20개의 자연 발생 아미노산뿐만 아니라 혼합물 또는 모호한 아미노산(핵산 표기법과 유사)을 나타낼 수 있다.
펩타이드는 직접 서열을 분석하거나 DNA 서열로부터 추론 될 수있다. 알려진 단백질 서열을 대조하는 큰 서열 데이터베이스가 존재한다.
| 아미노산 | 세 글자 약어 | 한 글자 약어 |
|---|---|---|
| 알라닌(Alanine) | Ala | A |
| 아르기닌(Arginine) | Arg | R |
| 아스파라긴(Asparagine) | Asn | N |
| 아스파르트산(Asparatic Acid) | Asp | D |
| 시스테인(Cysteine) | Cys | C |
| 글루탐산(Glutamic Acid) | Glu | E |
| 글루타민(Glutamine) | Gln | Q |
| 글라이신(Glycine) | Gly | G |
| 히스티딘(Histidine) | His | H |
| 아이소류신(isoleucine) | Ile | I |
| 류신(Leucine) | Leu | L |
| 라이신(Lysine) | Lys | K |
| 메티오닌(Methionine) | Met | M |
| 페닐알라닌(Phenylalanine) | Phe | F |
| 프롤린(Proline) | Pro | P |
| 세린(Serine) | Ser | S |
| 트레오닌(Threonine) | Thr | T |
| 트립토판(Tryptophan) | Trp | W |
| 티로신(Tyrosine) | Tyr | Y |
| 발린(Valine) | Val | V |
| 표기 | 설명 | 대표 잔기 |
|---|---|---|
| X | 모든 아미노산 또는 불명 | All |
| B | 아스파르트산 또는 아스파라긴 | D, N |
| Z | 글루탐산 또는 글루타민 | E, Q |
| J | 류신 또는 아이소류신 | I, L |
| Φ | 소수성 아미노산 | V, I, L, F, W, M |
| Ω | 방향성 아미노산 | F, W, Y, H |
| Ψ | 지방족 아미노산 | V, I, L, M |
| π | 분자량이 작은 아미노산 | P, G, A, S |
| ζ | 친수성 아미노산 | S, T, H, N, Q, E, D, K, R, Y |
| + | 양전하를 띠는 아미노산 | K, R, H |
| - | 음전하를 띠는 아미노산 | D, E |
변형
일반적으로 폴리펩타이드는 비분지형 중합체이므로 이들의 차 구조는 종종 그의 골격을 따라 아미노산 서열에 의해 특정된다. 그러나 단백질은 가장 일반적으로 이황화 결합에 의해 가교 결합 될 수 있으며, 단백질의 이황화결합에 관여하는 시스테인이 필요하다. 다른 가교에는 데스모신(Desmosine)이 포함된다.
이성질화
폴리펩타이드 사슬의 광학 이성질체는 라세미화를 겪을 수 있다. 순서는 변경되지 않지만 순서의 화학적 특성에 영향을 미친다. 특히 단백질에서 정상적으로 발견되는 L-아미노산은 자연적으로 이성질화 될 수있다. 대부분의 단백질 가수 분해 효소에 의해 분해 될 수 없는 D-아미노산을 형성하는 원자가 존재한다. 또한 프롤린은 펩타이드 결합에서 안정한 트랜스 이성질체를 형성 할 수 있다.

번역 후 변형
마지막으로 단백질은 다양한 번역 후 변형을 겪을 수 있다.
폴리펩타이드의 N 말단은 다음과 같이 변형 될 수 있다.
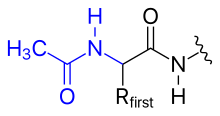
- 아세틸화

- 포르밀화

- 파이로 글루탐산(Pyroglutamate)
- 미리스토일화

폴리펩타이드의 C 말단은 다음과 같이 변형될 수 있다.
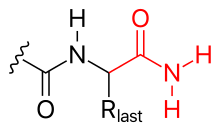
- 아미노화 (그림 참조)
- C 말단은 아미노화에 의해 차단 될 수 있다(따라서 음전하를 중화시킴).
- 글리코실포스파티딜이노시톨(GPI)의 부착
- 글리코실포스파티딜이노시톨(GPI)은 단백질을 세포막에 고정시키는 큰 소수성 인지질 그룹이다. 이는 아미노 결합을 통해 폴리펩타이드 C 말단에 부착되고, 이후 에탄올 아민에 연결되고, 최종적으로 포스파티딜이노시톨의 지질 부분에 연결된다.
마지막으로 펩타이드 곁사슬은 또한 공유적으로 변형 될 수 있다.
- 인산화
- 절단 이외에 인산화는 단백질의 가장 중요한 화학적 변형이다. 인산기는 세린, 트레오닌, 티로신 잔기의 곁사슬 하이드록시기에 부착 될 수 있으며, 그 부위에 음전하를 부가하고 비천연 아미노산을 생성한다. 이러한 반응은 인산화 효소에 의해 촉매되고 역반응은 인산 가수 분해 효소에 의해 촉매된다. 인산화 티로신은 종종 단백질이 서로 결합 할 수 있는 핸들로서 작용하는 반면, 세린/트레오닌의 인산화는 음전하 때문에 구조적 변화를 유도한다. 세린/트레오닌 잔기를 글루탐산으로 변화시킴으로써 세린/트레오닌의 인산화 효과를 확인할 수 있다.
- 당화
- 당화(Glycosylation)는 매우 일반적이고 이질적인 화학적 변형이 잘 일어난다. 당 Moiety는 세린/트레오닌의 곁사슬 하이드록시기 또는 아스파라긴의 곁사슬 아미노기에 부착 될 수있다. 이러한 부착물은 용해도 증가에서 복잡한 인식에 이르기까지 다양한 기능을 수행할 수 있다. 모든 당화는 튜니카마이신과 같은 특정 억제제로 차단 될 수 있다.
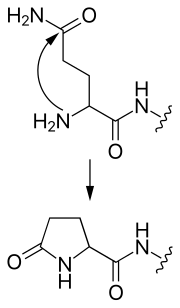
- 탈아미드화(석신이미드 형성)
- 이 변형에서, 아스파라긴 또는 아스파르트산의 곁사슬은 펩타이드 결합을 공격하여 대칭 숙신이미드 중간체를 형성한다. 중간체의 가수 분해는 아스파르트산 또는 β 아미노산, 동형 아스파르트산을 생성한다. 아스파라긴의 경우, 어느 하나의 생성물은 아미노기의 손실을 초래하여 탈아미드화가 된다.
- 하이드록시화
- 프롤린 잔기는 라이신(한 원자에서) 일 수 있는 것처럼, 두 원자 중 하나에서 하이드록시기를 가질 수 있다. 프롤린은 콜라겐의 중요한 성분으로, 결핍시 콜라겐이 불안정하다. 하이드록시화 반응은 비타민 C을 필요로 하는 효소에 의해 촉매되며, 결핍시 괴혈병과 같은 많은 결합 조직 질환을 유발한다.
- 메틸화
- 몇몇 단백질 잔기, 특히 라이신 및 아르기닌의 양전하 단백질이 메틸화 될 수 있다. 아르기닌 잔기는 핵산-인산 골격과 상호 작용하고 단백질-DNA 복합체에서 염기 잔기, 특히 구아닌과 수소 결합을 형성한다. 라이신 잔기는 단일 결합, 이중 결합, 삼중 결합으로 메틸화 될 수 있다. 그러나 메틸화는 곁사슬 상의 양전하를 변화시키지 않는다.
- 아세틸화
- 라이신 아미노기의 아세틸화는 N 말단의 아세틸화와 화학적으로 유사하다. 그러나 기능적으로, 라이신 잔기의 아세틸화는 핵산에 대한 단백질의 결합을 조절하는데 사용된다. 라이신 상의 양전하의 제거는(음전하) 핵산에 대한 정전기적 인력을 약화시킨다.
- 황화
- 티로신은 원자로 황화 반응이 일어난다. 다소 특이하게도 이러한 변형은 소포체가 아닌 골지체에서 발생한다. 인산화 티로신과 유사하게, 황화 티로신은 세포 표면상의 케모카인 수용체에서 특정 인식을 위해 사용된다. 인산화와 마찬가지로, 황화는 이전 중립 위치에 음전하를 추가한다.

- 프레닐화 및 팔미토일화

- 소수성 이소프렌(파네실, 제라닐, 제라닐제라닐기) 및 팔미토일기는 세포막에 단백질을 고정시키는 시스테인 잔기의 원자이다. GPI 및 미리토일앵커와 달리, 이들 그룹은 반드시 말단에 추가 될 필요는 없다.

- 카복실화
- 글루탐산 곁사슬에 여분의 글루탐산 그룹 (이중 음전하)을 첨가하여 글루타민 잔기를 생성하는데, 비교적 드문 변형이다. 이것은 칼슘과 같은 경질 금속 이온에 대한 결합을 강화하는 데 사용된다.
- ADP-리보실화
- 큰 ADP-리보실기는 이종 효과를 가지면서 단백질 내의 여러 유형의 곁사슬로 옮길 수 있다. 이 변형은 Vibrio cholerae, Corynebacterium diphtheriae, Bordetella pertussis와 같은 이종 세균의 강력한 독소를 표적으로 한다.
- 유비퀴틴화 및 SUMO화
- 다양한 형태의 접힌 단백질이 그들의 C 말단에서 다른 단백질의 라이신의 곁사슬 암모늄기에 부착 될 수 있다. 유비퀴틴이 이들 중 가장 흔하며, 일반적으로 유비퀴틴 표시된 단백질이 분해되어야 한다는 신호를 보낸다.
위에서 열거된 폴리펩타이드 변형은 진핵생물의 세포 내 소기관인 소포체에서 발생하고, 리보솜에서 합성 된 후에 발생한다.
많은 다른 화학 반응(시아닐화)은 생물학적 시스템에서는 발견되지 않지만 화학자들에 의해 단백질에 적용되었다.
분열 및 결찰
상기에 열거된 것 이외에, 단백질의 1차 구조의 가장 중요한 변형은(가수 분해 또는 단백질 가수 분해 효소에 의한)펩타이드 절단이다. 단백질은 종종 비활성 전구체 형태로 합성된다. 일반적으로 N 말단 또는 C 말단 세그먼트는 단백질의 활성 자리를 차단하여 그 기능을 억제한다. 단백질은 억제 펩타이드를 절단함으로써 활성화된다.
일부 단백질은 스스로를 쪼개는 힘을 가지고 있다. 전형적으로 세린의 하이드록시기(드물게 트레오닌) 또는 시스테인 잔기의 티올기는 앞선 펩타이드 결합의 카르보닐 탄소를 공격하여 사면 결합 된 중간체(히드록시옥사졸리딘, 세린/트레오닌으로 분류 됨) 또는 히드록시티아졸리딘(시스테인중간체)가 그 예이다. 아미드 형태는 일반적으로 자유에너지에 의해 선호되므로, 이 중간체는 아미드 형태로 되돌아가는 경향이 있다. 그러나 추가적인 분자 상호 작용은 아미드 형태를 덜 안정적으로 만들 수 있다. 대신에 아미노기가 방출되어 펩타이드 결합 대신에 에스터 결합(세린/트레오닌) 또는 티오에스터 결합(시스테인)이 생성된다. 이 화학 반응을 'N-O 아실기 이동'이라고 한다.
- 간단한 가수 분해는 치환된 아미노기가 새로운 N 말단이 되는 폴리펩타이드 사슬을 분할한다. 이것은 glycosylasparaginase의 성숙에서 볼 수 있다. 또한 β 제거 반응은 사슬을 분할하지만, 새로운 N 말단에서 피루보일기를 초래한다. 이 피루보일기는 일부 효소, 특히 피루보일기의 전자 흡인력을 이용하는 S-DC( S-adenosylmethionine decarboxylase)와 같은 탈탄산효소(decarboxylaase)에서 공유 결합된 촉매 보조인자로 사용될 수 있다. 분자 내 에스터 교환으로 분지형 폴리펩타이드가 생성된다. 인테인(Intein)에서, 새로운 에스터 결합은 곧 C 말단 아스파라긴에 의한 분자 내 공격에 의해 파괴된다. 분자 간 에스터 교환 반응은 헤지호그 단백질 처리에서 볼 수 있듯이, 전체 세그먼트를 한 폴리펩타이드에서 다른 폴리펩타이드로 옮길 수 있다.
다른 분자의 1차 구조
임의의 선형 사슬 이형 중합체는 단백질에 대한 용어의 사용과 유사하게 1차 구조를 가질 수 있지만, 이 사용은 단백질과 관련하여 매우 일반적인 사용과 비교하여 드물다. 광범위한 2차 구조를 갖는 RNA에서, 염기의 선형 사슬은 일반적으로 DNA에서와 같이 서열로 지칭된다. 다당류와 같은 다른 생물학적 중합체도 기본 구조는 아니지만 1차 구조를 갖는 것으로 간주 될 수 있다.
2차 및 3차 구조와의 관계
생물학적 중합체의 1차 구조는 대부분 3차원 형태(3차 구조)를 결정한다. 단백질 서열은 2차 구조의 세그먼트 또는 막 관통 영역과 같은 국소 특징을 예측하는데 사용될 수 있다. 그러나 단백질 접힘의 복잡성은 현재 그 서열만으로 단백질의 3차 구조를 예측하는 것을 금지한다. 유사한 상동성 서열의 구조(동일한 단백질 군의 구성원)를 알면 상동성 모델링에 의해 3차 구조를 매우 정확하게 예측할 수 있다. 전장 단백질 서열이 이용 가능하다면, 등전점과 같은 일반적인 생물리학적 특성 을 추정 할 수 있다.