

감수분열(meiosis, ![]() i/maɪˈoʊsɪs/, 감소하는 분열이라는 점에서 그리스어로 줄어든다는 의미의 μείωσις, meiosis에서 유래함)이란 유성 생식을 하는 생물들이 정자나 난자 같은 생식자(gamete)를 생성하기 위해 종자세포(germ cell)에서 수행하는 특별한 세포 분열 방법이다. 감수분열은 총 2회의 분열을 하며, 각 염색체 사본을 하나씩(홑배수체; n)만 가지는 4개의 딸세포를 만든다. 또한, 분열하기 전에 아버지와 어머니로부터 받은 각 염색체의 유전 물질은 교차를 통해 섞여서 각 염색체는 새로운 조합을 형성한다. 이후 아버지와 어머니 각각이 감수분열을 통해 홑배수체 세포를 만들면, 이 세포는 수정이라는 과정을 통해 서로 융합하여 다시 각 염색체 사본을 2개씩 가지는 두배수체(2n) 접합자(zygote)를 만든다.
i/maɪˈoʊsɪs/, 감소하는 분열이라는 점에서 그리스어로 줄어든다는 의미의 μείωσις, meiosis에서 유래함)이란 유성 생식을 하는 생물들이 정자나 난자 같은 생식자(gamete)를 생성하기 위해 종자세포(germ cell)에서 수행하는 특별한 세포 분열 방법이다. 감수분열은 총 2회의 분열을 하며, 각 염색체 사본을 하나씩(홑배수체; n)만 가지는 4개의 딸세포를 만든다. 또한, 분열하기 전에 아버지와 어머니로부터 받은 각 염색체의 유전 물질은 교차를 통해 섞여서 각 염색체는 새로운 조합을 형성한다. 이후 아버지와 어머니 각각이 감수분열을 통해 홑배수체 세포를 만들면, 이 세포는 수정이라는 과정을 통해 서로 융합하여 다시 각 염색체 사본을 2개씩 가지는 두배수체(2n) 접합자(zygote)를 만든다.
감수분열에서 오류가 발생하면 비정배수체(aneuploidy; 비정상적인 염색체 수) 생식자가 만들어진다. 이런 생식자가 수정을 하여 접합자를 형성하면, 이 접합자는 제대로 발생하지 못하고 유산되거나, 대부분의 경우 발생 장애를 갖고 태어난다.
DNA 복제가 먼저 일어난 뒤 감수분열을 진행하며, 감수분열 결과 생긴 4개의 딸세포에서는 원래 모세포보다 염색체 수가 절반으로 줄어든다. 두 번 진행되는 분열은 각각 순서대로 감수분열 I(감수 1분열)과 감수분열 II(감수 2분열)라고 한다. 감수분열을 시작하기 전 S기 때, 각 염색체의 DNA가 복제되어 2개의 동일한 자매 염색분체를 가진다. 자매 염색분체는 코히신이라는 단백질로 서로 붙들려 있다. 감수분열 이전의 S기를 '감수분열이전 S기'나 '감수분열 S기'라고도 부른다. DNA 복제가 끝나자마자 감수분열할 세포는 G2기와 비슷하면서도 좀 더 긴 감수분열 전기에 돌입한다. 이때 상동 염색체 쌍은 유전자 재조합이라는 프로그래밍된 과정을 겪으면서 DNA가 잘리고 다시 복구된다. 유전자 재조합 과정을 통해 염색체 쌍끼리 유전 정보를 서로 교환한다. 재조합이 일어나면 염색분체끼리 교차(crossover)가 이루어지고, 이때 생겨나는 물리적인 교차 지점을 교차점(단수: chiasma, 복수: chiasmata; 그리스 문자 Chi(X)에서 유래함)이라고 한다. 교차는 대부분의 생물에서 각 상동 염새체 쌍이 감수분열 I 동안 서로 멀리 떨어져 분리될 수 있게 도와준다. 감수분열 I 이후 만들어지는 딸세포는 모세포의 절반만큼의 염색체 수를 가지는 홑배수체 세포이다.
감수분열 II 동안에 자매 염색분체를 서로 붙들고 있던 코히신 단백질이 방출되면서 체세포분열이 일어날 때와 마찬가지로 자매 염색분체가 서로 분리된다. 일부 경우에는 감수분열로 만들어진 딸세포 4개 모두가 정자, 홀씨(포자), 꽃가루 같은 생식자(gamete)를 형성한다. 그러나 암컷 동물은 일반적으로 감수분열로 만들어진 딸세포 4개 중 3개를 극체로 만들어 제거하고, 오직 딸세포 1개만 난자로 발달한다. 감수분열 동안 염색체 수가 절반으로 줄어들기 때문에 각 부모가 만든 생식자는 수정을 통해 융합하여 각 염색체를 2세트씩 가지는 두배수체 접합자를 형성한다. 이렇게 감수분열과 수정을 번갈아 진행하면서 유성 생식이 이루어지며, 세대를 거치면서도 똑같은 수의 염색체를 유지한다. 사람으로 예를 들어 보면, 두배수체 사람 세포는 1쌍의 성염색체를 포함하여 총 23쌍의 염색체를 가지는데(총 46개), 그중 절반은 어머니로부터, 나머지 절반은 아버지로부터 물려받은 것이다. 여기서 감수분열을 하면 염색체 1세트(23개)를 포함하는 홑배수체 생식자(난자나 정자)가 만들어진다. 2개의 생식자(난자와 정자)가 융합하면, 이때 만들어지는 접합자는 다시 어머니와 아버지로부터 염색체 23개씩을 물려받아 두배수체가 된다. 감수분열을 하는 모든 생명체는 서로 염색체 수가 다를지라도 똑같은 패턴으로 생활주기를 이어간다.
감수분열은 동물, 식물, 진균을 포함하여 유성 생식을 하는 모든 단세포 생물과 다세포 생물(다 진핵생물)에서 일어난다. 감수분열 과정은 난자발생과 정자발생을 할 때 반드시 거쳐야 하는 과정이다.
개요
감수분열 과정은 일반적인 세포 분열 과정인 체세포분열과 관련되어 있지만, 이 둘은 크게 2가지 관점에서 서로 차이가 난다.
| 재조합 | 감수분열 | 각 쌍의 염색체 2개(각 부모로부터 물려받은 염색체) 사이에서 유전자를 서로 섞어서 모든 생식자가 고유의 유전적 조합을 가지는 재조합 염색체를 만든다. | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 체세포분열 | DNA 손상 복구가 필요한 경우에만 일어난다;
일반적으로 동일한 자매 염색분체 사이에서 재조합이 일어나기 때문에 유전자 변화가 나타나지 않는다. |
||||||||||
| 염색체 수 (배수성) |
감수분열 | 유전적으로 서로 다른 4개의 딸세포를 생성하며, 각 딸세포는 모세포 염색체의 절반만큼의 염색체를 가진다. | |||||||||
| 체세포분열 | 유전적으로 똑같은 2개의 딸세포를 생성하며, 각 딸세포는 모세포 염색체와 똑같은 수의 염색체를 가진다. | ||||||||||
감수분열은 각 염색체 사본을 2개씩 가지는, 즉 상동 염색체를 가지는 두배수체 세포에서 시작한다. 먼저 세포는 DNA 복제 과정을 거치고, 이로써 각 상동 염색체는 동일한 자매 염색분체 2개로 이루어진다. 이후 각 상동 염색체 세트는 상동 재조합을 통해 유전 물질을 서로 교환하며, 이때 종종 상동 염색체 쌍 사이에 물리적인 연결(교차)이 나타난다. 첫 번째 감수분열에서 방추장치에 의해 상동 염색체가 분리된다. 그 이후 세포는 DNA 복제를 추가로 하지 않고 바로 두 번째 분열을 진행한다. 이때 자매 염색분체가 분리되면서 딸세포가 만들어지며, 총 4개의 홑배수체 세포가 생성된다. 암컷 동물은 이 패턴에서 살짝 다른 방법을 사용하여 감수분열 결과 큰 난자 1개와 2~3개의 작은 극체를 생성한다. 재조합으로 인해 각 염색분체는 어머니와 아버지의 유전 정보가 서로 섞인 새로운 조합을 포함할 수 있기 때문에 자손은 부모와 유전적으로 다르다. 뿐만 아니라 각 생식자는 재조합이 일어나지 않은 온전한 어머니 염색체, 온전한 아버지 염색체, 재조합 염색체 조합으로 이루어질 수도 있다. 이렇게 유성 생식으로부터 형성되는 유전적 다양성으로 형질의 다양성이 나타나며, 이는 자연 선택이 작동할 수 있는 원동력 중 하나다.
진핵생물에서 하나의 세포가 2개의 동일한 딸세포로 분열하는 체세포분열의 메커니즘은 감수분열에서도 많이 이용된다. 일부 식물과 진균, 원생생물의 감수분열은 홀씨(포자; spore)를 만드는데, 홀씨는 수정 없이도 스스로 분열하면서 성장할 수 있는 홑배수체 세포이다. 브델로이데아 목(Bdelloidea)과 같은 일부 진핵생물은 감수분열을 수행할 능력이 없는 대신 처녀생식을 통해 생식할 능력을 획득하였다.
감수분열은 아키아나 박테리아에서는 일어나지 않으며, 이들은 일반적으로 이분법을 통해 무성 생식한다. 그러나 수평 유전자 전달이라는 과정을 통해 '유성' 과정을 수행할 수도 있는데, 이 경우 한 박테리아나 아키아로부터 다른 박테리아나 아키아로 DNA가 전달되고, 서로 다른 세포에서 유래한 DNA 분자가 재조합된다.
역사
1876년 독일의 생물학자 오스카 헤트비히(Oscar Hertwig)가 처음으로 성게 알에서 감수분열을 발견하고 묘사했다. 1883년 벨기에 동물학자 에두아 바 브네든(Edouard Van Beneden)은 아스카리스 속(Ascaris) 회충 알에서 감수분열을 염색체 수준으로 또 한 번 묘사했다. 그러나 오직 1890년 독일의 생물학자 아우구스트 베이즈만(August Weismann)이 생식과 유전 측면에서 감수분열의 중요성을 묘사했는데, 베이즈만은 만약 염색체 수가 유지되어야 한다면 두배수체 세포 1개가 홑배수체 세포 4개로 바뀌기 위해서 2번의 세포 분열이 필요하다고 언급했다. 1911년 미국 유전학자 토머스 헌트 모건(Thomas Hunt Morgan)은 초파리 드로소필라 멜라노가스터(Drosophila melanogaster)의 감수분열 과정에서 교차를 발견했으며, 이는 유전 형질이 염색체에서 전달된다는 사실을 확립해주었다.
'meiosis'라는 용어는 '줄어듦'을 뜻하는 그리스어 μείωσις에서 유래하였다. J.B. 파머(J.B. Farmer)와 J.E.S. 무어(J.E.S. Moore)는 1905년에 이 용어를 'maiosis'라고 독특하게 번역하여 처음으로 생명과학 분야에서 사용하기 시작했다.
플레밍이 '헤테로타입'과 '호모타입'이라고 정의한 2번의 분열에서 나타나는 핵 변화의 전체 과정을 아울러 우리는 Maiosis나 Maiotic phase라는 용어를 사용할 것을 제안한다.
이후 코니케(Koernicke; 1905), 그리고 판텔과 드 시네티(Pantel and De Sinety; 1906)는 그리스어 표기 원칙에 따라 철자를 'meiosis'라고 바꾸었다.
일본은 20세기 초반 'meiosis'의 어원을 살려 '수가 줄어드는 세포 분열'이라는 의미에서 '減數分裂'(감수분열)이라는 한자어로 번역했고, 일제 강점기와 그 이후 우리나라가 이 한자어를 그대로 음독하여 표기함으로써 '감수분열'이라는 용어가 만들어졌다.
현재 생명과학 교과서를 위해 교육부가 편찬하는 생명과학 편수 자료에는 meiosis를 '감수분열'과 '생식세포 분열' 2가지 용어로 제시하고 있다. 하지만, '생식세포(reproductive cell)'는 생명과학 용어가 아닌 데다가, 같은 편수 자료를 보면 '생식세포'는 별개의 용어인 'gamete', 'germ cell', 'generative cell' 모두를 지칭하는 잘못 쓰이는 용어이다. 따라서 현재 '생식세포 분열'이라는 표현을 쓰는 교과서는 원래대로 '감수분열'이라고 고쳐 표기하고 사용해야 한다.
교육부에서 지정한 '감수 1분열'과 '감수 2분열'이라는 용어도 모두 '減数第一分裂'(감수제일분열)과 '減数第二分裂'(감수제이분열)이라는 일본식 표현을 따른 잘못된 표기이므로, 학계에서 사용하는 '감수분열 I'과 '감수분열 II'라는 표현을 사용할 것을 권고한다.
시기
감수분열은 감수분열 I과 감수분열 II로 나뉜다. 또 각각의 분열은 핵분열 I과 세포질분열 I, 그리고 핵분열 II와 세포질분열 II로 나뉜다. 감수분열을 준비하는 단계는 체세포분열 주기에서의 사이기와 패턴이 똑같다.사이기는 아래와 같이 3단계로 시기로 나뉜다.
- 성장1(G1)기: 이 시기는 매우 활발한 시기로, 세포는 성장에 필요한 효소와 구조 단백질 등의 어마어마한 단백질을 합성한다. G1기에서 각 염색체는 선형 DNA 분자를 하나씩 포함한다.
- 복제(S)기: 유전 물질이 복제된다. 각 염색체는 복제되어서 중심절에서 서로 붙어 있는 2개의 동일한 자매 염색분체로 이루어진다. 염색체 수가 일정하게 유지되기 때문에 염색체가 복제되더라도 세포의 배수성은 변하지 않는다. 또한 염색체는 광학 현미경에서 보일 정도로 빽빽하게 응축하지 않는다. 응축 과정은 감수분열의 전기 I 동안 일어난다.
- 성장2(G2)기: 체세포분열과 다르게 감수분열에는 G2기가 없다. 감수분열 전기 I이 체세포분열에서의 G2기와 거의 비슷하다.
사이기 이후 감수분열 I이 진행되고, 그 이후 감수분열 II가 진행된다. 감수분열 I 때는 복제된 상동 염색체가 분리되어 각 딸세포 2개로 이동하고, 각 염색체는 여전히 2개의 자매 염색분체로 이루어진다. 따라서 이때 염색체 수가 절반으로 감소한다. 감수분열 II 동안에는 자매 염색분체끼리 분리되어 딸염색체가 4개의 딸세포로 각각 나뉘어 들어간다. 두배수체 생물의 경우 감수분열로부터 만들어진 딸세포는 홑배수체이고, 각 염색체 사본은 1개씩만 포함한다. 일부 종들은 감수분열 I과 감수분열 II 사이에 중간기(interkinesis)라는 휴식기를 거친다.
감수분열 I과 II는 체세포분열의 세포 주기와 비슷하게 각각 전기, 중기, 후기, 말기 단계로 나뉜다. 따라서 감수분열은 감수분열 I(전기 I, 중기 I, 후기 I, 말기 I)과 감수분열 II(전기 II, 중기 II, 후기 II, 말기 II)로 구성된다.

감수분열 과정 동안 특정 유전자에서 훨씬 많은 전사가 이루어진다. 감수분열 단계에 특이적인 mRNA 발현이 강하게 나타나는 것과 더불어, 전반적인 번역 조절(예를 들어 미리 형성된 mRNA의 선택적 사용)도 일어나기 때문에 궁극적으로 감수분열 시기 동안 감수분열 단계에 특이적인 단백질 발현을 조절한다. 이렇게 하여 전사 조절과 번역 조절 모두가 감수분열을 수행하는 데 적절하게끔 세포를 전체적으로 다시 구조화한다.
감수분열 I
감수분열 I 때 2가 염색체(2n, 4c)를 형성하고 있던 상동 염색체가 분리된다. 그 결과 염색분체 쌍을 각각 가지는 2개의 홑배수체 세포(1n, 2c)가 만들어진다. 배수성이 두배수체에서 홑배수체로 줄어들기 때문에 감수분열 I은 감소하는 분열이라고도 불린다. 감수분열 II는 자매 염색분체가 분리되어 4개의 홑배수체 딸세포(1n, 1c)를 만들기 때문에 체세포분열과 마찬가지로 균등 분열이다.


전기 I
전기 I은 감수분열에서 가장 긴 시기이다(총 14일 동안 진행되는 생쥐의 감수분열 중 전기 I은 13일 동안 지속된다). 전기 I 동안 아버지와 어머니로부터 물려받은 상동 염색체 쌍이 서로 이어지고(synapsis), 유전 정보가 교환되며(상동 재조합으로), 염색체 1쌍당 최소 1개 이상의 교차가 형성된다. 이때 교차점(chiasma)을 통해 교차를 관찰할 수 있다. 교차 과정은 상동 염색체가 안정한 쌍을 형성할 수 있도록 해주며, 이에 따라 첫 감수분열 때 염색체가 정확하게 분리될 수 있도록 해준다. 각 부모로부터 유래하여 복제된 뒤 서로 쌍을 이룬 염색체를 2가 염색체(bivalent; 염색체 2개) 또는 4분 염색체(tetrad; 염색분체 4개)라고 부른다. 전기 I은 염색체의 모양에 따라 가는섬유기, 접합기, 굵은섬유기, 겹섬유기, 이동기의 5단계로 세분화된다.
가는섬유기 (Leptotene)
전기 I의 첫 단계는 그리스어로 '가는 실'이라는 뜻의 가는섬유기(leptonema; 세사기)이다. 전기 I의 가는섬유기 때 2개의 복제된 자매 염색분체로 이루어진 각각의 염색체는 '개별화'되어 핵에서 관찰할 수 있는 가닥을 형성한다. 각 염색체는 코히신에 의해 고리가 선형으로 배열되는 형태를 띤다. 그리고 이음실복합체(synaptonemal complex)의 가쪽 요소(lateral element)가 모인 뒤, 고리가 달리는 '축 요소(axial element)'를 형성한다. 이 단계 때 Spo11이라는 효소에 의해 재조합이 시작되면서 프로그래밍된 이중나선 절단이 만들어진다(생쥐의 경우 감수분열당 약 300회). 이 과정으로 RAD51과 DMC1로 덥힌 단일가닥 DNA가 만들어지며, 이 단일가닥 DNA는 상동 염색체를 파고들어 염색체 축 사이에 다리를 형성하고, 이에 따라 상동 염색체가 짝을 이뤄 함께 정렬된다(생쥐의 경우 ~400 nm 거리에서).
접합기 (Zygotene)
가는섬유기 다음으로는 그리스어로 '짝지은 실'이라는 뜻의 접합기(zygotene)가 시작된다. 텔로미어가 핵의 한쪽 끝에 모여 있는 모여 있는 일부 생물의 경우 염색체 모양 때문에 '꽃다발 단계(bouquet stage)'로 불리기도 한다. 접합기에서는 이음실복합체에 가로 요소(transverse element)와 중심 요소(central element)가 부착하면서 상동 염색체가 훨씬 더 가까워지고(~100 nm) 안정하게 짝을 이룬다(이 과정을 연접이라고 함). 연접(synapsis)은 지퍼와 비슷한 방식으로 재조합 결절에서 시작해서 형성될 것이라고 여겨진다. 짝지은 염색체는 2가 염색체나 4분 염색체라고 불린다.
굵은섬유기 (Pachytene)
그리스어로 '굵은 실'이라는 뜻의 굵은섬유기(pachytene /ˈpækɪtiːn/ PAK-i-teen; 태사기, 후사기) 단계는 모든 상염색체가 서로 이어져 있는 단계이다. 굵은섬유기 때 교차가 일어난 상동 염색체는 가는섬유기 때 나타났던 이중가닥 절단의 복구가 완료된다. 대부분의 DNA 절단은 교차를 형성하지 않은 채 복구되기 때문에 유전자 전환이 이루어지지 않는다. 그러나 DNA 이중가닥 절단 중 일부(염색체당 최소 1개 이상)는 비자매(상동) 염색체 사이에서 교차를 형성하여 유전 정보를 교환시킨다. 성염색체의 경우 전체적으로 동일하지 않기 때문에, 거짓상염색체 영역이라고 불리는 서로 상동인 작은 영역에서만 유전 정보를 교환한다. 상동 염색분체 사이에서의 유전 정보 교환의 결과 유전 정보의 재조합이 일어나고, 각 염색체는 재조합 결과 형성된 공백 없이 이전에 가졌던 것과 같은 유전 정보의 완전한 집합을 가진다. 염색체는 이음실복합체와 구별되지 않기 때문에 실제로 평범한 광학 현미경으로는 교차가 어떻게 일어나는지 관찰할 수 없고, 교차점도 다음 단계 전까지 보이지 않는다.
겹섬유기 (Diplotene)
그리스어로 '2개의 실'이라는 뜻의 겹섬유기(diplotene; 쌍사기) 동안 이음실복합체가 해체하고, 상동 염색체가 서로 약간 분리된다. 그러나 각 2가 염색체를 구성하는 상동 염색체는 교차점에서 단단히 결합한 상태로 존재한다. 교차점은 상동 염색체가 세포의 반대편 극으로 이동하는 것이 허용되는 후기 I로 전환되기 전까지는 염색체에 남아 있다.
사람 태아의 난자발생의 경우, 발생하고 있는 모든 난모세포는 겹섬유기까지 발달한 다음 태어나기 전까지 전기 I에 정지해 있는다. 이렇게 중단된 단계를 연장기(dictyate)라고 부른다. 사춘기 이후에 난모세포의 배란이 준비되어 감수분열이 다시 시작되기 전까지 연장기가 계속 이어진다.
이동기 (Diakinesis)
그리스어로 '이동함'이라는 뜻의 이동기 단계에서는 염색체가 더 응축한다. 이 시기가 감수분열에서 처음으로 4개의 염색분체가 실제로 관찰되는 시기이다. 교차가 이루어진 지점은 서로 얽히며 효과적으로 겹쳐져 교차점이 뚜렷하게 관찰되게 해준다. 이런 점을 제외하면, 이 단계의 나머지는 체세포분열의 전중기와 비슷하다. 핵이 사라지고, 핵막은 소포로 해체되며, 감수분열 방추가 형성된다.
감수분열 방추 형성
체세포분열을 하는 세포와 다르게, 사람과 생쥐 난모세포는 감수분열 방추를 만드는 중심체를 가지지 않는다. 생쥐의 경우 대략 80개의 미세관형성중심(MTOC)이 난모세포질 안에서 한 영역을 형성한 뒤 미세관을 응집시키기 시작하고, 이 미세관은 염색체까지 뻗어 방추부착점(kinetochore)에 부착한다. 시간이 지나면서 MTOC는 두 극이 형성되기 전까지 합쳐져 배럴 모양의 방추를 만들어 낸다. 사람 난모세포의 경우 방추 미세관 응집이 염색체에서 시작된 뒤, 최종적으로 염색체를 둘러싸도록 커지면서 별 모양을 형성한다. 이후 염색체는 방추부착점이 미세관과 부착하는 지점인 방추의 적도를 향하여 미세관을 따라 미끄러지듯이 움직인다.
중기 I
상동 염색체 쌍은 함께 중기판으로 이동한다. 세포 양쪽의 방추 극으로부터 뻗어 나온 방추부착점 미세관은 상동 염색체 쌍 각각의 방추부착점에 결합하고, 이후 상동 염색체 쌍은 방추를 이등분하면서 적도판을 따라 배열된다. 이때 계속해서 상동 염색체의 두 방추부착점에 연결된 미세관에 의해 2가 염색체에 균형을 잡아주는 힘이 가해진다. 이런 부착을 두극 부착(bipolar attachment)이라고 한다. 동일한 적도판을 따라 2가 염색체들이 배열될 때 각 2가 염색체는 중기판에 무작위한 방향으로 배열되는데, 이는 멘델이 밝힌 독립 유전의 원리를 설명하는 물리적 근거이다. 단백질 복합체인 코히신은 염색체가 복제되는 순간부터 후기 I까지 자매 염색분체를 함께 붙들어둔다. 체세포분열의 경우 방추부착점 미세관이 양쪽 방향으로 염색체를 잡아당길 때 장력이 발생한다. 세포는 이런 장력을 감지하여 모든 염색체가 적절한 방향으로 배열되기 전까지는 후기를 진행시키지 않는다. 감수분열의 경우 장력을 발생시키기 위해 자매 염색분체를 잇는 코히신과 더불어 염색체 1쌍당 최소 1개 이상의 교차가 필요하다(염색체 분리 참고).
후기 I
방추부착점 미세관이 짧아지면서 상동 염색체(각각 자매 염색분체 쌍으로 이루어짐)를 세포 반대편 극으로 끌어당긴다. 극성 미세관은 길어지면서 중심체를 더 멀리 밀어내고, 그 결과 세포는 중심으로부터 길어진다. 모든 코히신이 분해되는 체세포분열과 다르게, 염색체 팔에 결합된 코히신만 분해되고, 중심절을 둘러싸는 코히신은 슈고신(Shugoshin; '수호신'의 일본어)이라는 단백질에 의해 보호 받으며 그대로 남아 있다. 이 때문에 자매 염색분체는 상동 염색체가 분리되는 순간에 함께 붙어 있을 수 있다.
말기 I
첫 번째 감수분열은 사실상 염색체가 양쪽 극에 도달하면서 끝난다. 이제 각 딸세포는가 가지는 염색체 수는 절반으로 줄어들었지만, 각 염색체는 염색분체 쌍으로 이루어져 있다. 방추 네트워크를 이루던 미세관이 사라지고, 새로운 핵막이 각 홑배수체 염색체 세트를 둘러싸며 생성된다. 염색체는 다시 염색질로 풀린다. 그러나 경우에 따라 핵막은 다시 형성되지 않을 수도 있고, 염색체도 사이기만큼 완전히 풀리지 않는다.
동물 세포의 경우 핵막을 집으면서, 식물 세포의 경우 세포벽을 형성하면서 세포질분열이 일어나고, 그 결과 2개의 딸세포 형성이 완료된다. 그러나 세포질분열이 완전히 완료되지 않으면 딸세포끼리 '세포질다리'로 연결되면서 감수분열 II가 끝날 때까지 딸세포는 서로 세포질을 공유한다. 자매 염색분체는 말기 I 동안에도 붙어 있다.
세포는 휴식하는 시기인 중간기(interkinesis; 사이기 II라고도 부름)에 돌입할 수도 있다. 이 시기에는 DNA 복제가 일어나지 않는다.
감수분열 II
감수분열 II는 두 번째 감수분열로, 자매 염색분체가 분리된다. 이 과정은 체세포분열과 메커니즘이 비슷하지만, 유전적 결과는 근본적으로 다르다. 감수분열 II가 일어나면 감수분열 I에서 만들어졌던 2개의 홑배수체 세포(자매 염색분체 2개를 포함하는 n 염색체)로부터 총 4개의 홑배수체 세포(n 염색체)가 만들어진다. 감수분열 II의 단계는 전기 II, 중기 II, 후기 II, 말기 II이다.
전기 II에서는 핵소체와 핵막이 다시 사라지고, 염색체는 응축하여 짧고 굵어진다. 중심체는 세포 양극으로 이동하고, 2번째 감수분열을 하기 위해 방추 섬유를 배열한다.
중기 II에서는 각 중심체로부터 뻗어 나온 방추 섬유가 각 염색체의 방추부착점에 각각 부착한다. 새로 형성되는 적도판은 감수분열 I의 적도판과 비교했을 때 90°회전된 상태이다.
후기 II에서는 중심절 주변에 남아 있던 코히신이 더 이상 슈고신의 보호를 받지 않음에 따라 쪼개지고, 자매 염색분체가 분리된다. 이때 자매 염색분체였던 것은 이제 딸염색체가 되어 서로 반대편 극을 향하여 이동한다.
말기 II에서 감수분열 II가 끝나며, 이때 염색체는 풀리면서 길어지고 방추는 해체된다. 핵막이 다시 형성되고, 최종적으로 세포질분열이 일어나 홑배수체 세트 염색체를 가지는 총 4개의 딸세포가 생성된다.
감수분열은 이제 완료되었고 새로운 딸세포 4개가 만들어지면서 끝난다.
기원과 기능
감수분열의 기원과 기능은 현재 과학적으로 잘 밝혀져 있지 않지만, 이는 진핵생물에서 유성 생식의 진화에 관해 근본적인 통찰력을 제공할 것이다. 어떻게 진핵생물의 진화 과정에서 성이 나타나게 되었는지, 유성 생식이 어떤 기본적인 기능을 하는지, 왜 유성 생식이 유지되는지에 관해 아직은 생물학자 사이에서 의견 일치가 없다. 그러나 유성 생식은 12억 년 전에 진화해 생겼고, 식물, 진균, 동물을 포함하여 원래부터 유성 생식을 했던 종의 후손은 거의 모두 유성 생식을 한다는 사실은 명백하다.
감수분열은 진핵생물의 유성 생식 주기에서 나타나는 중요한 사건이다. 감수분열은 각각 모세포에 비해 절반만큼의 염색체를 가지는 홑배수체 세포(생식자)를 만들어내는 생활 주기의 한 단계이다. 보통 서로 다른 개체로부터 만들어진 2개의 홑배수체 생식자가 수정이라는 과정을 통해 융합하면 성 주기가 완성된다.
감수분열은 진핵생물 전체에 걸쳐 공통적이다. 이는 효모 같은 단세포 생물에서도 일어나고, 사람과 같은 다세포 생물에서도 일어난다. 진핵생물은 약 22억 년 전에 원핵생물로부터 만들어졌고, 초기의 진핵생물은 단세포 생물과 비슷했다. 진핵생물에서 성을 이해하기 위해서는 (1) 어떻게 감수분열이 단세포로 된 진핵생물로부터 생겨났는지, (2) 감수분열의 기능이 무엇인지 알 필요가 있다.
감수분열 동안 DNA가 새롭게 조합되는 것은 돌연변이와 함께 유전적 다양성을 일으키는 중요한 근원이며, 그 결과 이득이 되는 대립유전자의 새로운 조합이 만들어진다. 감수분열은 두 가지 방법으로 생식자의 유전적 다양성을 만든다. 첫째, 멘델의 독립유전의 원리이다. 중기 I 때 중기판을 따라 상동 염색체 쌍들이 서로 독립적인 방향으로 배열되고 중기 II 때도 자매 염색분체들이 서로 독립적인 방향으로 배열되는데, 이는 각 딸세포에게(궁극적으로 생식자에게) 무작위적이고 독립적으로 유전 정보가 들어갈 수 있도록 해준다. 둘째, 교차이다. 전기 I 동안 상동 재조합으로 인해 상동 염색체 영역이 물리적으로 교환되어 염색체에서 새로운 조합의 유전 정보가 만들어진다.
전기 I 정지
암컷 포유류와 새들은 난모세포가 미래의 배란에 필요한 모든 것들을 포함한 채로 태어나며, 이 난모세포는 감수분열 전기 I에 정지된 상태에 있다. 예를 들어서 사람 여성의 경우, 난모세포는 임신 3~4달 사이 태아 시기 때 형성되며, 태어날 때부터 이미 가지고 있다. 이렇게 전기 I이 정지된 단계(연장기; dictyate)는 수십 년 동안 지속될 수 있으며, 4개의 지놈 사본이 난모세포에 존재하고 있다. 이렇게 난모세포가 지놈 사본이 4개인 상태로 정지되어 있으면, 여분의 유전 정보를 사용해서 생식계 DNA에 손상이 일어났을 때 복구할 수 있다. 이 복구 과정은 상동 재조합 복구 메커니즘으로 나타난다. 전기 I 때 정지된 난모세포는 외부 요인에 의한 이중가닥 절단과 같은 DNA 손상을 효율적으로 복구할 수 있는 능력이 매우 뛰어나다. DNA 복구 능력은 여성 생식계에서 중요한 질 조절 메커니즘으로 작동하고, 여성의 생식 능력을 결정한다.
나타남
생활 주기에서


감수분열은 유성 생식과 관련하여 진핵생물의 생활 주기에서 감수분열과 수정 과정을 순환하면서 나타난다. 이는 일반적인 체세포분열과도 함께 일어난다. 다세포 생물의 경우, 생명체가 성장할 때 두배수체와 홑배수체 전환 사이에 중간 단계가 존재한다. 생활 주기의 특정 단계에서 종자세포는 생식자를 만든다. 체세포는 생명체의 몸을 구성하고, 생식자 생성에는 관여하지 않는다.
홑배수체 상태와 두배수체 상태가 교대로 나타나면서 감수분열과 수정이 주기적으로 반복된다. 생물은 생활 주기의 대부분을 두배수체 상태로 보낼 수도 있고(두배수체 생활 주기), 홑배수체 상태로 보낼 수도 있고(홑배수체 생활 주기), 둘 다로 보낼 수도 있다(홑두배수체 생활 주기, 여기에는 홑배수체 상태와 두배수체 상태의 2가지 구별되는 시기가 존재함). 이 관점으로 보면, 유성 생식을 이용하는 생활 주기는 생물 시기의 위치에 따라 세 종류로 구별된다.
두배수체 생활 주기(diplontic life cycle)(생식자 만들기 전에 감수분열)의 경우, 생물체는 두배수체이고, 접합자(zygote)라고 불리는 두배수체 세포로부터 성장한다. 사람도 두배수체 생활 주기로 살아간다. 생물의 두배수체 생식계 줄기세포는 감수분열을 통해 홑배수체 생식자(수컷은 정자, 암컷은 난자)를 만들며, 생식자는 수정하여 접합자를 형성한다. 두배수체 접합체는 다시 체세포분열을 반복하면서 생명체로 성장한다.
홑배수체 생활 주기(haplontic life cycle)(접합자 만든 이후 감수분열)의 경우, 생명체는 홑배수체이고, 생식자(gamete)라고 불리는 하나의 홑배수체 세포가 증식하고 분화함으로써 발생한다. 서로 다른 성을 가진 두 생명체에서 나온 홑배수체 생식자는 두배수체 접합자를 형성한다. 접합자는 만들어진 즉시 감수분열을 하여 4개의 홑배수체 세포를 생성한다. 홑배수체 생식자는 다시 체세포분열을 하며 성장해서 하나의 생명체를 만든다. 많은 진균과 많은 원생동물이 홑배수체 생활 주기로 살아간다.
마지막으로 홑두배수체 생활 주기(haplodiplontic life cycle)(홀씨 감수분열이나 중간 감수분열)의 경우, 생명체는 홑배수체 상태와 두배수체 상태를 서로 교대한다. 결과적으로 이 주기는 세대 교대라고 알려져 있다. 두배수체 생물의 생식계 세포는 감수분열을 통해 홑배수체 홀씨(spore)를 만든다. 홀씨는 체세포분열을 통해 증식해서 홑배수체 생명체로 성장한다. 이후 홑배수체 생명체의 생식자가 다른 홑배수체 생명체의 생식자와 서로 합쳐지면 다시 두배수체 생명체가 된다. 이처럼 홑두배수체 생활 주기는 두배수체 생활 주기와 홑배수체 생활 주기의 융합으로 여겨질 수 있다.
식물과 동물에서
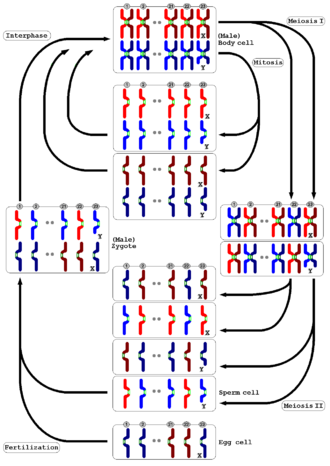
감수분열은 모든 동물과 식물에서 일어난다. 동물과 식물 둘 다 모세포의 절반만큼 염색체 수를 가지는 생식자가 만들어진다는 점은 똑같지만, 상세한 과정은 서로 다르다. 동물은 감수분열을 통해 생식자를 직접 만든다. 그러나 육상 식물과 일부 조류(algae)는 두배수체 홀씨체(sporophyte; 포자체) 세대에서 감수분열이 일어나 홑배수체 홀씨를 생성하는 세대 교대가 일어난다. 이 홀씨는 체세포분열을 통해 수가 늘어나며 홑배수체 배우체로 발생한다. 배우체는 추가적인 감수분열 없이 바로 생식자를 만든다. 동물과 식물 둘 다 생식자의 최종 단계는 융합해서 다시 염색체 수를 원래대로 회복하는 것이다.
포유류에서
암컷의 경우 난모세포(oocyte)라는 세포에서 감수분열이 일어난다. 각 일차난모세포는 감수분열을 통해 두 번 나뉘는데, 각 분열마다 불균등하게 나뉜다. 첫 번째 분열에서는 딸세포 하나와 그보다 훨씬 작은 극체 하나가 만들어지는데, 극체는 두 번째 분열을 진행할 수도 있고 안 할 수도 있다. 감수분열 II에서 딸세포가 두 번째로 분열하면서 두 번째 극체와 하나의 홑배수체 세포를 생성하는데, 이 세포는 거대해져서 난자(ovum)가 된다. 그러므로 암컷의 일차 난모세포가 감수분열을 하면 하나의 성숙한 난자와, 1~2개의 극체가 만들어진다.
암컷은 감수분열 시기 중간에 감수분열이 중단된다. 성숙한 난모세포는 감수분열 I의 전기 I 때 정지해 있고, 난포세포(follicle)라는 체세포 보호 껍질에 싸여 휴면 상태에 놓인다. 각 월경 주기가 시작될 때, 난포자극호르몬이 뇌하수체 앞엽에서 분비되어 약간의 난포를 난포발생(folliculogenesis)이라는 과정으로 성숙하게 만든다. 이 과정 동안 난모세포는 감수분열을 다시 시작하여 감수분열 II의 중기 II까지 계속한 뒤, 배란 직전에 중기 II에서 다시 멈춘다. 사람의 난포발생 과정 동안 주로 하나의 난포만 선택되어 발생하고, 나머지 난포는 폐쇄된다. 암컷의 감수분열은 난자발생(oogenesis)이라는 과정을 거쳐 진행되는데, 연장기(dictyate) 단계라고 알려진 긴 시간 동안 감수분열이 중단된다는 점과, 중심체 도움 없이 진행된다는 점에서 일반적인 감수분열과 다르다.
수컷은 고환의 정세관에서 정자발생(spermatogenesis) 과정을 통해 감수분열이 일어난다. 정자발생 동안 일어나는 감수분열은 정모세포(spermatocyte라는 특정 종류의 세포에서만 일어나며, 정모세포는 성숙하여 정자(en:spermatozoa)가 된다. 원시종자세포의 감수분열은 암컷보다 한참 늦은 사춘기 때 일어난다. 사춘기 이전까지 수컷의 고환 조직은 감수분열을 촉진하는 물질인 레티노산을 분해시킴으로써 감수분열을 억제한다. 사춘기가 되면 정세관에 있는 버팀세포(세톨리 세포; sertoli cell)가 스스로 레티노산을 만들기 시작하면서 감수분열이 진행된다. 레티노산에 대한 감수성은 나노스(nanos)와 DAZL라고 불리는 단백질에 의해서도 조절된다. 레티노산 생산 효소의 유전자 기능상실 연구를 통해 레티노산은 출생 이후 정조세포의 분화를 촉진하여 정모세포가 정자가 되기 위해 필요하다는 사실이 밝혀졌지만, 레티노산은 감수분열이 시작되는 동안 필요한 것은 아니다.
암컷 포유류는 원시종자세포가 배아의 난소로 이동한 이후 즉시 감수분열을 시작한다. 일부 연구는 원시 콩팥(중간콩팥)에서 유래한 레티노산이 암컷 배아 난소에 있는 난조세포의 분화를 촉진시키고, 수컷 배아 정소 조직은 레티노산을 분해함으로써 감수분열을 억제한다는 사실을 밝혔다. 그러나 레티노산 생산 효소의 유전자 기능상실 연구는 레티노산이 배아 발생 동안 진행되는 암컷 감수분열이나 출생 이후 시작되는 수컷 감수분열 둘 다에 필요하지 않다는 사실을 밝혔다.
편모류
진핵생물 대부분은 감수분열 동안 총 2번 분열한다(때때로는 교차 없이). 그러나 바퀴벌레 Cryptocercus의 장에 서식하는 파라바살리드류(parabasalid)나 옥시모나드류(parabasalid) 같은 일부 편모류(flagellate)는 감수분열 동안 총 1번 분열하는 매우 드문 감수분열을 진행한다.
사람 유전과 질병에서의 역할
사람 염색체 23쌍에서 일어나는 재조합은 실제 염색체를 다시 분배할 것뿐만 아니라, 각 염색체의 부분도 다시 분배한다. 남성에 비해 여성에서 1.6배 더 많은 재조합이 일어난다고 추정된다. 또한, 평균적으로 여성의 재조합은 중심절 근처에서 많이 일어나고, 남성의 재조합은 텔로미어에서 많이 일어난다. 평균 100만 bp(1 Mbp)는 1 cMorgan (cm = 1% 재조합 빈도)과 맞먹는다. 이는 100만 bp당 1% 확률로 재조합이 일어난다는 뜻이다. 교차 빈도는 아직 불확실하다. 효모, 생쥐, 사람에서는 감수분열 하는 세포당 200회 이상의 이중가닥 절단(DSB)이 일어난다고 추정된다. 그러나, DSB 중 일부(생물에 따라 ~5-30%)만 교차를 형성하는데, 그 결과 사람의 경우 염색체당 1-2개의 교차만 형성된다.
비분리
감수분열 I에서 염색체가 정상적으로 나뉘거나, 감수분열 II에서 자매 염색분체가 나뉘는 현상을 분리(disjunction)라고 한다. 그러나 분리가 정상적으로 일어나지 않을 경우는 비분리(nondisjunction)라고 부른다. 비분리 결과 생식자는 특정 염색체 수보다 더 많거나 더 적은 염색체를 가지게 되고, 이는 세염색체증이나 홑염색체증을 일으키는 흔한 메커니즘이다. 비분리는 세포 생식 시기인 감수분열 I이나 감수분열 II 때 일어날 수도 있고, 체세포분열 동안 일어날 수도 있다.
대부분 홑염색체증이나 세염색체증 사람 배아는 생존하지 못한다. 그러나 21번 염색체처럼 작은 염색체의 세염색체증과 같은 일부 비정배수성은 생존할 수 있다. 이런 비정배수성의 표현형은 심각한 발생에서부터 무증상까지 다양하다. 의학적으로 아래와 같은 예를 정의하고 있지만, 다른 예들도 얼마든지 많다.
- 다운 증후군 - 21번 염색체의 세염색체증
- 파타우 증후군 - 13번 염색체의 세염색체증
- 에드워드 증후군 - 18번 염색체의 세염색체증
- 클라인펠터 증후군 - 남성에서 X 염색체가 추가로 있음 - XXY, XXXY, XXXXY 등
- 터너 증후군 - 여성에서 X 염색체가 하나 없음 - XO
- 트리플 X 증후군 - 여성에서 X 염색체가 추가로 있음
- 제이콥스 증후군 - 남성에서 Y 염색체가 추가로 있음
사람 난모세포에서 비분리가 일어날 가능성이 여성의 나이가 증가함에 따라 커지는데, 이는 시간이 지나면서 코히신이 손실되기 때문이다.
체세포분열과 비교
감수분열을 이해하기 위해 체세포분열과 비교해보는 것이 도움 된다. 아래 표는 감수분열과 체세포분열의 차이점을 보여준다.
| 감수분열 | 체세포분열 | |
|---|---|---|
| 최종 결과 | 일반적으로 4개의 딸세포, 각각 모세포 염색체 수의 절반을 가짐 | 2개의 딸세포, 모세포와 염색체 수가 똑같음 |
| 기능 | 두배수체 생활주기를 가지고 유성 생식을 하는 진핵생물에서 생식자를 생성함 | 세포 증식, 성장, 재생, 무성 생식 |
| 일어나는 위치 | 거의 모든 진핵생물 (동물, 식물, 진균, 원생생물); 생식샘에서, 생식자가 되기 전에(두배수체 생활주기); 접합자가 된 이후에(홑배수체 생활주기); 홀씨가 되기 전에(홑두배수체 생활주기) |
모든 진핵생물의 모든 증식하는 세포에서 |
| 단계 | 전기 I, 중기 I, 후기 I, 말기 I, 전기 II, 중기 II, 후기 II, 말기 II |
전기, 전중기, 중기, 후기, 말기 |
| 모세포와 유전적으로 동일한가? | 아니오 | 예 |
| 교차가 일어나는가? | 예, 일반적으로 상동 염색체의 각 쌍 사이에서 일어남 | 매우 드묾 |
| 상동 염색체가 짝을 이루는가? | 예 | 아니오 |
| 세포질분열 | 말기 I과 말기 II 때 일어남 | 말기 때 일어남 |
| 중심절 분리 | 후기 I 때 안 일어나고 후기 II 때 일어남 | 후기 때 일어남 |
분자 조절
감수분열을 하는 세포가 어떻게 이를 진행하는지는 아직 잘 알려져 있지 않다. 성숙촉진인자(maturation promoting factor)는 개구리 난모세포의 감수분열 때 특정 역할을 할 것으로 보인다. 진균 종류 중 하나인 Schizosaccharomyces pombe에서는 MeiRNA가 부착한 단백질이 감수분열을 시작하도록 해준다.
중심절 영역인 CDE1에 부착하는 효모의 CEP1 유전자 산물이 감수분열 I 동안 염색체가 짝지어 줄 것이라고 제안되었다.
감수분열 재조합은 이중가닥 절단에 의해 매개되며, 이는 Spo11 단백질에 의해 촉매된다. 또한 Mre11, Sae2, Exo1 단백질도 DNA 절단과 재조합을 일으킨다. DNA 절단이 일어난 이후 상동인 부분에서 재조합이 일어난다. 재조합은 이중 할리데이 이음(dHJ) 경로나, 합성 의존적 가닥 어닐링(SDSA) 방법 중 하나로 이루어진다(SDSA 방법에서는 교차가 일어나지 않는다).
보아하니 감수분열에도 체크포인트가 존재한다. Schizosaccharomyces pombe에서는 Rad 단백질과 Mek1(FHA 카이네이스 도메인 포함), Cdc25, Cdc2 단백질, 그리고 아직 알려지지 않은 인자들이 체크포인트를 형성할 것으로 생각된다.
척추동물의 난자발생에서는 세포정지인자(CSF; cytostatic factor)가 감수분열 II로 전환시키는 역할을 한다.
더 보기
참고 문헌
인용한 책
- Freeman S (2005). 《Biological Science》 3판. Upper Saddle River, NJ: Pearson Prentice Hall. ISBN 9780131409415.